Anti-Inflammatory Activity of Spearmint (Mentha spicata) Essential Oil in Human Dermal Fibroblasts
*Corresponding Author(s):
Xuesheng HanDoterra International, LLC, Pleasant Grove, Utah, United States
Tel:+1 8014377976,
Email:lhan@doterra.com
Tory L Parker
Doterra International, LLC, Pleasant Grov, Utah, United States
Tel:+1 8014377957,
Email:tparker@doterra.com
Abstract
The use of Spearmint (Mentha spicata) Essential Oil (SEO) in skin care products has become popular. In this study, we evaluated the impact of a commercially available SEO on 17 protein biomarkers that perform critical functions in inflammation and tissue remodeling in a validated human dermal fibroblast system, which was designed to model the pathology of chronic inflammation. The impact of SEO on genome-wide gene expression was also evaluated. SEO showed robust antiproliferative effects in diseased human skin cells and significantly inhibited the increased production of two pro-inflammatory biomarkers Vascular Cell Adhesion Molecule-1 (VCAM-1) and interferon-inducible T-cell α chemoattractant (I-TAC). In contrast, SEO significantly increased the levels of Monocyte Chemoattractant Protein-1 (MCP-1), Epidermal Growth Factor Receptor (EGFR), Matrix Metalloproteinase-1 (MMP-1) and Tissue Inhibitor of Metalloproteinase-1 (TIMP-1). SEO significantly modulated global gene expression and altered signaling pathways, many of which are critical in the inflammatory and tissue remodeling processes. Further analysis showed that the gene expression data obtained were consistent with the observed anti-inflammatory and tissue remodeling effects of SEO. This study provided the first evidence of SEO’s anti-inflammatory activity in human dermal fibroblasts, which suggested that SEO might be a promising candidate for anti-inflammatory skincare products.
Keywords
INTRODUCTION
Spearmint (Mentha spicata) Essential Oil (SEO) is rich in carvone and limonene and has a refreshing aroma. In comparison with peppermint essential oil, SEO is perceived to be a milder option for topical application in individuals with sensitive skin. This could be one of the reasons for the increased popularity of SEO in skin care, beauty and healthcare products. Studies in various models have demonstrated its therapeutic potential, which include antiproliferative, antibacteria l, antifungal, anticonvulsant and antiemetic activities [1-7]. However, studies on the effects of SEO in human skin cells are scarce. Although the anti-inflammatory properties of limonene, a major active component of SEO, have been demonstrated in both pre-clinical and clinical models; to our knowledge, no studies have been conducted to evaluate the anti-inflammatory activity of SEO in human dermal fibroblasts.
We investigated the biological activity of a commercially available SEO in a validated human dermal fibroblast cell culture line designed to model the pathology of chronic inflammation [8,9]. We analyzed the impact of SEO on 17 important protein biomarkers closely related to the inflammatory and tissue remodeling pathways. We also studied the regulatory effect of SEO on genome-wide gene expression.
MATERIALS AND METHODS
Cell cultures
Protein based readouts
The quantitative biomarker data were presented as the mean log10 relative expression level (compared with the respective mean vehicle control value) ± Standard Deviation (SD) of triplicate measurements. The differences in biomarker levels between the SEO and vehicle treated cultures were tested for significance with an unpaired Student’s t-test. A p-value < 0.05, outside of the significance envelope, with an effect size of at least 10% (more than 0.05 log10 ratio units), was considered statistically significant.
RNA isolation
Microarray analysis for genome-wide gene expression
The raw data were uploaded into and analyzed for quality control metrics using the bead array package [13,14]. The data were normalized using quantile normalization and then re-annotated and filtered to remove probes that were non-specific or mapped to intronic or intragenic regions [15,16]. The remaining probe sets comprised the data set for the rest of the analysis. Fold-change expression for each value was calculated as the log2 ratio of SEO to the vehicle control. These fold change values were uploaded into the Ingenuity Pathway Analysis (IPA, Qiagen, Redwood City, CA, USA) to generate the networks and pathway analyses.
Reagents
The chemical composition of the oil was analyzed using Gas Chromatography-Mass Spectrometry (GC-MS). Detailed methods of GC-MS have been previously described [12]. GC-MS analysis of SEO indicated that its major chemical constitutes (i.e., >5%) are carvone (68%) and limonene (19%). The GC-MS chromatography of SEO has been included in supplementary figure 1.
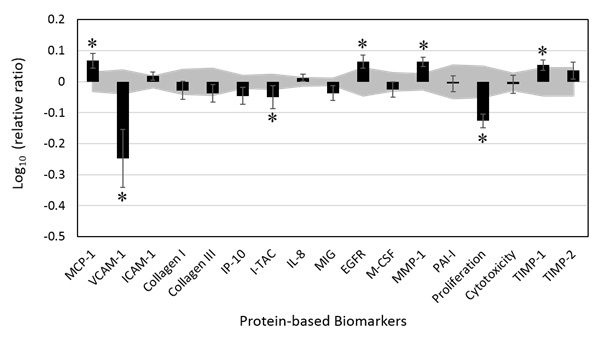 Figure 1: Bioactivity profile of spearmint essential oil (SEO, 0.011% v/v) in a human dermal fibroblast system (HDF3CGF).
Figure 1: Bioactivity profile of spearmint essential oil (SEO, 0.011% v/v) in a human dermal fibroblast system (HDF3CGF).*indicates a biomarker designated with “key activity”; the biomarker value for the treated cells was significantly different (p<0.05) from that of the vehicle control at the tested concentration, with an effect size of at least 10% (more than 0.05 log ratio units).
MCP-1 - Monocyte Chemoattractant Protein 1
VCAM-1 - Vascular Cell Adhesion Molecule 1
ICAM-1 - Intracellular Cell Adhesion Molecule 1
IP-10 - Interferon Gamma-Induced Protein 10
I-TAC - Interferon-inducible T-cell Alpha Chemoattractant
IL-8 - Interleukin 8
MIG - Monokine Induced by Gamma Interferon
EGFR - Epidermal Growth Factor Receptor
M-CSF - Macrophage Colony-Stimulating Factor
MMP-1 - Matrix Metalloproteinase 1
PAI-1 - Plasminogen Activator Inhibitor 1
SRB - Sulforhodamine B
TIMP - Tissue Inhibitors of Metalloproteinase
RESULTS AND DISCUSSION
Bioactivity profile of SEO in the human dermal fibroblast system HDF3CGF
Overall, SEO demonstrated diverse effects on multiple protein molecules, including significant antiproliferative activity in dermal fibroblasts. In addition, SEO significantly suppressed the increased production of two pro-inflammatory biomarkers, Vascular Cell Adhesion Molecule-1 (VCAM-1) and Interferon-inducible T-cell Alpha Chemoattractant (I-TAC). In contrast, SEO significantly increased the level of Monocyte Chemoattractant Protein 1 (MCP-1), which is also a pro-inflammatory chemokine. The important biomarkers related to tissue remodeling, namely Epidermal Growth Factor Receptor (EGFR), Matrix Metalloproteinase-1 (MMP-1) and Tissue Inhibitor of Metalloproteinase-1 (TIMP-1) were also significantly increased by SEO. SEO had no significant effect on the other biomarkers tested in the experimental system.
Zhao et al., reported that SEO reduced pulmonary inflammation in rats with chronic obstructive pulmonary disease [17]. de Sousa et al., observed the spasmolytic activity of carvone and limonene, which can assist proper respiratory and cardiovascular functioning [18]. A recent human study demonstrated the beneficial effects of SEO on lung function and exercise performance [19], which may be at least partially attributed to its anti-inflammatory and spasmolytic feature. Fitsiou et al., recently showed that SEO rich in carvone (85.4%) exhibited antiproliferative activity in cancer cells [1].
Carvone and limonene were found to modulate immune response in BALB/c mice [20]. Studies on carvone derivatives also demonstrated their anti-inflammatory potential [18]. de Sousa et al., reported that orally administered hydroxydihydrocarvone exerted anti-inflammatory effects in rats and mice [21,22]. Marques et al. studied the anti-inflammatory activity of cyane-carvone, a synthetic derivative of carvone, and found that it significantly decreased IL-1β and TNF-α levels and significantly inhibited paw edema in mice [22].
Several studies of limonene showed that it demonstrated anti-inflammatory potential in a variety of disease models, possibly through the suppression of pro-inflammatory cytokines [23,24], p38 mitogen activated protein kinase, nuclear factor-κB, c-Jun N-terminal kinase , and extracellular signal-regulated kinase [25-28]. Notably, Rufino et al. observed that limonene increased the gene expression of TIMP-1 and decreased the expression of MMP-1 in a mouse model of osteoarthritis [23]. The results of the current study showed that SEO significantly increased the protein levels of TIMP-1 and MMP-1 in a human skin disease model.
Studies have also shown evidence that carvone and limonene act as transdermal permeation enhancers [29]. Limonene has been found to promote skin repair and inhibit skin tumorigenesis [30,28] .Both these properties are partly attributable to its anti-inflammatory activity. These features may further contribute to the beneficial effects of SEO on skin health.
Effects of SEO on genome-wide gene expression in the HDF3CGF system
Further, IPA studies indicated that the pattern of SEO bioactivity matched multiple canonical pathways listed in the literature-validated signaling pathway database (Figure 2). Many of these pathways play critical roles in the processes of inflammation, tissue remodeling, and immune modulation. The overall inhibitory effect of SEO on these genes and signaling pathways appeared consistent with its observed anti-inflammatory properties; the Supplementary Material contains more detailed information.
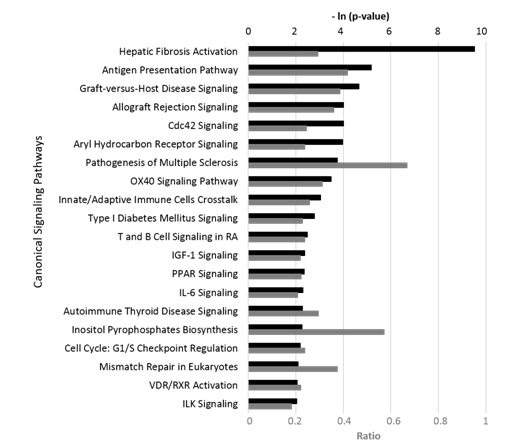
OX40 - Tumor necrosis factor receptor super family, member 4
RA - Rheumatoid Arthritis
IGF-1 - Insulin-Like Growth Factor 1
PPAR - Peroxisome Proliferator-Activated Receptors
VDR - Vitamin D Receptor
RXR - Retinoid X Receptor
ILK - Integrin-Linked Kinase
Collectively, these data showed that SEO, of which the main components are carvone and limonene, had significant antiproliferative and anti-inflammatory activities in human dermal fibroblasts. In addition, these findings suggested that SEO might be able to modulate immune responses during the wound healing process and promote improved wound healing. The results of the current study were consistent with those of existing studies, which indicated the therapeutic potential of SEO as an anti-inflammatory candidate for skin care products. Further research into its biological mechanism of action in specific disease models, as well as its clinical efficacy and safety, is recommended.
CONCLUSION
To our knowledge, this is the first study to evaluate the biological activity of SEO in pre-inflamed human dermal fibroblast cells. SEO showed significant antiproliferative activity through the inhibition of VCAM-1and I-TAC levels and the increased expression of MCP-1, EGFR, MMP-1 and TIMP-1. Genome-wide gene expression analysis demonstrated that SEO exerted a robust and diverse impact on the expression of many genes. Many of the genes and pathways highly affected by SEO were critically involved in the inflammatory and tissue remodeling processes. These results were consistent with those of other studies that reported the anti-inflammatory and wound healing potential of SEO. Although the clinical efficacy and safety of SEO administration are still undetermined, our study indicated that SEO was a promising candidate for use in anti-inflammatory skin products.
ACKNOWLEDGMENT
This study was funded by d?TERRA (Pleasant Grove, UT, USA) and conducted at DiscoverX (Fremont, CA, USA). Xuesheng Han interpreted the data and drafted the manuscript. Cody Beaumont developed and conducted the GC-MS assays. Tory Parker oversaw the study design, methodology, and study agent and model selection. All authors reviewed and approved the final manuscript.
CONFLICTS OF INTEREST
Xuesheng Han and Cody Beaumont and Tory Parker are employees of d?TERRA, the manufacturer of the study agent (SEO).
REFERENCES
- Fitsiou E, Mitropoulou G, Spyridopoulou K, Tiptiri-Kourpeti A, Vamvakias M, et al. (2016) Phytochemical profile and evaluation of the biological activities of essential oils derived from the greek aromatic plant species Ocimum basilicum, Mentha spicata, Pimpinella anisum and Fortunella margarita. Molecules 21.
- Hussain AI, Anwar F, Nigam PS, Ashraf M, Gilani AH (2010) Seasonal variation in content, chemical composition and antimicrobial and cytotoxic activities of essential oils from four Mentha s Journal of the Science of Food and Agriculture 90: 1827-1836.
- Shahbazi Y (2015) Chemical Composition and In Vitro Antibacterial Activity of Mentha spicataEssential Oil against Common Food-Borne Pathogenic Bacteria. Journal of Pathogens 916305: 5.
- Houicher A, Hechachna H, Teldji H, Ozogul F (2016) In Vitro Study of the Antifungal Activity of Essential Oils Obtained from Mentha spicata, Thymus vulgaris, and Laurus nobilis. Recent Pat Food Nutr Agric 8: 99-106.
- Nardoni S, Giovanelli S, Pistelli L, Mugnaini L, Profili G, et al. (2015) In Vitro Activity of Twenty Commercially Available, Plant-Derived Essential Oils against Selected Dermatophyte Species. Nat Prod Commun 10: 1473-1478.
- Koutroumanidou E, Kimbaris A, Kortsaris A, Bezirtzoglo E, Polissio M, et al. (2013) Increased seizure latency and decreased severity of pentylenetetrazol-induced seizures in mice after essential oil administration. Epilepsy Research and Treatment 532657: 6.
- Tayarani-Najaran Z, Talasaz-Firoozi E, Nasiri R, Jalali N, Hassanzadeh MK (2013) Antiemetic activity of volatile oil from Mentha spicata and Mentha × Piperita in chemotherapy-induced nausea and vomiting. Ecancermedicalscience 7: 290.
- Kunkel EJ, Dea M, Ebens A, Hytopoulos E, Melrose J, et al. (2004) An integrative biology approach for analysis of drug action in models of human vascular inflammation. FASEB J 18: 1279-1281.
- Bergamini G, Bell K, Shimamura S, Werner T, Cansfield A, et al. (2012) A selective inhibitor reveals PI3Kγdependence of TH17 cell differentiation. Nature Chemical Biology 8: 576-582.
- Berg EL, Yang J, Melrose J, Nguyen D, Privat S, et al. (2010) Chemical target and pathway toxicity mechanisms defined in primary human cell systems. Journal of Pharmacological and Toxicological Methods 61: 3-15.
- Han X, Parker TL (2017) Anti-inflammatory, tissue remodeling, immunomodulatory, and anticancer activities of oregano (Origanum vulgare) essential oil in a human skin disease model. Biochimie Open 4: 73-77.
- Han X, Beaumont C, Stevens N (2017) Chemical composition analysis and in vitrobiological activities of ten essential oils in human skin cells. Biochimie Open 5: 1-7.
- R Development Core Team (2011) R: A Language and Environment for Statistical Computing. The R Foundation for Statistical Computing, Vienna, Austria.
- Dunning MJ, Smith ML, Ritchie ME, Tavaré S (2007) beadarray: R classes and methods for Illumina bead-based data. Bioinformatics 23: 2183-2184.
- Bolstad BM, Irizarry RA, Astrand M, Speed TP (2003) A comparison of normalization methods for high density oligonucleotide array data based on variance and bias. Bioinformatics 19: 185-193.
- Barbosa-Morais NL, Dunning MJ, Samarajiwa SA, Darot JFJ, Ritchie ME, et al. (2010) A re-annotation pipeline for Illumina BeadArrays: improving the interpretation of gene expression data. Nucleic Acids Res 38:
- Zhao CZ, Wang Y, Tang FD, Zhao XJ, Xu QP, et al. (2008) [Effect of Spearmint oil on inflammation, oxidative alteration and Nrf2 expression in lung tissue of COPD rats]. Zhejiang Da Xue Xue Bao Yi Xue Ban 37: 357-363.
- de Sousa DP, Mesquita RF, de Araújo Ribeiro LA, de Lima JT (2015) Spasmolytic Activity of Carvone and Limonene Enantiomers. Nat Prod Commun 10: 1893-1896.
- Jaradat NA, Al Zabadi H, Rahhal B, Hussein AM, Mahmoud JS, et al. (2016) The effect of inhalation of Citrus sinensis flowers and Mentha spicata leave essential oils on lung function and exercise performance: a quasi-experimental uncontrolled before-and-after study. J Int Soc Sports Nutr 13: 36.
- Raphael TJ, Kuttan G (2003) Immunomodulatory activity of naturally occurring monoterpenes carvone, limonene, and perillic acid. Immunopharmacol Immunotoxicol 25: 285-294.
- de Sousa DP, Camargo EA, Oliveira FS, de Almeida RN (2010) Anti-inflammatory activity of hydroxydihydrocarvone. Z Naturforsch C 65: 543-550.
- Marques TH, Marques ML, Medeiros JV, Silva RO, dos Reis Barbosa AL, et al. (2014) Cyane-carvone, a synthetic derivative of carvone, inhibits inflammatory response by reducing cytokine production and oxidative stress and shows antinociceptive effect in mice. Inflammation 37: 966-977.
- Rufino AT, Ribeiro M, Sousa C, Judas F, Salgueiro L, et al. (2015) Evaluation of the anti-inflammatory, anti-catabolic and pro-anabolic effects of E-caryophyllene, myrcene and limonene in a cell model of osteoarthritis. Eur J Pharmacol 750: 141-150.
- Yoon WJ, Lee NH, Hyun CG (2010) Limonene suppresses lipopolysaccharide-induced production of nitric oxide, prostaglandin E2, and pro-inflammatory cytokines in RAW 264.7 macrophages. J Oleo Sci 59: 415-421.
- Chi G, Wei M, Xie X, Soromou LW, Liu F, et al. (2013) Suppression of MAPK and NF-κB pathways by limonene contributes to attenuation of lipopolysaccharide-induced inflammatory responses in acute lung injury. Inflammation 36: 501-511.
- Rehman MU, Tahir M, Khan AQ, Khan R, Oday-O-Hamiza, et al. (2014) D-limonene suppresses doxorubicin-induced oxidative stress and inflammation via repression of COX-2, iNOS, and NFκB in kidneys of Wistar rats. Exp Biol Med (Maywood) 239: 465-476.
- Kim K-N, Ko Y-J, Yang H-M, Ham Y-M, Roh SW, et al. (2013) Anti-inflammatory effect of essential oil and its constituents from fingered citron (Citrus medica L. var. sarcodactylis) through blocking JNK, ERK and NF-κB signaling pathways in LPS-activated RAW 264.7 cells. Food and Chemical Toxicology 57: 126-131.
- Chaudhary SC, Siddiqui MS, Athar M, Alam MS (2012) D-Limonene modulates inflammation, oxidative stress and Ras-ERK pathway to inhibit murine skin tumorigenesis. Human & Experimental Toxicology 31: 798-811.
- Krishnaiah YS, Raju V, Shiva Kumar M, Rama B, Raghumurthy V, et al. (2008) Studies on optimizing in vitro transdermal permeation of ondansetron hydrochloride using nerodilol, carvone, and limonene as penetration enhancers. Pharm Dev Technol 13: 177-185.
- d'Alessio PA, Mirshahi M, Bisson JF, Bene MC (2014) Skin repair properties of d-Limonene and perillyl alcohol in murine models. Antiinflamm Antiallergy Agents Med Chem 13: 29-35.
SUPPLEMENTARY FILE
Citation: Han X, Beaumont C, Parker TL (2017) Anti-Inflammatory Activity of Spearmint (Mentha spicata) Essential Oil in Human Dermal Fibroblasts. J Cytol Tissue Biol 4: 015.
Copyright: © 2017 Xuesheng Han, et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.