Antibodies against Desmoglein 1, Desmoglein 3 and BP180 in Healthy Tunisians Blood Donors without Immunopathological Phenotype
*Corresponding Author(s):
Amina ToumiDepartment Of Medical And Health Sciences, Habib Bourguiba Hospital, Emirates College Of Technology, Abu Dhabi, United Arab Emirates
Tel:+21674451660,
Email:at_toumi@yahoo.fr
Abstract
Pemphigus Vulgaris (PV), Pemphigus Foliaceus (PF) and Bullous Pemphigoid (BP) are a group of rare organ-specific diseases that affect skin and mucous membranes. Each subtype of pemphigus has its own anti-Desmogleins (Dsgs) autoantibodies profile: Dsg1 is the auto-antigen in PF whereas Dsg3 is the in PV.
We identified 2 PF patients with positive levels of anti-Dsg3 Abs and also 4 healthy blood donors with high levels of anti-Dsg1 Abs and anti-Dsg3 Abs.
The purpose of this study was to examine epitopes recognized by anti-Dsg1/anti-Dsg3 Abs and to identify potential autoreactive HLAII-restricted helper T cell epitopes shared among Dsg1+/Dsg3+ subjects that could explain the simultaneous occurrence of these auto-Abs.
On IIF, only one PF patients had antibodies to the keratinocyte surface. Interestingly, the ELISA for BP180 was also positive in 2 healthy blood donors. IP-IB analysis showed immunoreactivity to more than one EC domain of Dsg1 and towards the C-terminal EC5 domain of Dsg3. MHC II gene analysis demonstrated that all healthy blood donors had at least one protective allele associated to PF.
We confirm in this study that the onset and course of this disease depend on a variable interaction between predisposing and inducing genetic, environmental, hormonal, and immunological factors.
Keywords
ABBREVIATIONS
PF: Pemphigus Foliaceus;
PV: Pemphigus Vulgaris;
BP: Bullous Pemphigoid,
Dsg: Desmoglein,
HLA: Human Leucocyte Antigen
INTRODUCTION
Autoimmune mucocutaneous blistering diseases such as Pemphigus Vulgaris (PV), Pemphigus Foliaceus (PF), Bullous Pemphigoid (BP), and Mucous Membrane Pemphigoid (MMP), are a group of rare organ-specific diseases that affect skin and multiple mucous membranes [1,3]. There is increasing evidence that the clinical phenotype is strongly related to the antibody profile. Desmogleins (Dsgs) are the target antigens in pemphigus [3,4]. Each subtype has its own anti-Dsg autoantibodies profile: Dsg1 is the auto-antigen in PF whereas Dsg3 is the autoantigen in PV [1].
In discordance with the above, we identified a group of 2 PF patients with positive levels of anti-Dsg3 Abs and also 4 healthy blood donors with positive levels of both anti-Dsg1 Abs and anti-Dsg3 Abs (Table 1). Indeed, it has been well documented that autoimmune diseases may coexist in the same patient, either sequentially or concurrently [5-7]. Anti-Dsg3 autoantibodies are detected in the sera of a small number of PF patients (<7%) [8]. Moreover, it has been reported that multiple keratinocyte cadherins such as Dsg1, Dsg2, Dsg3, Dsg4, Dsc1, Dsc2, Dsc3, and E-cadherin, are recognized by healthy controls in endemic area as well as PF patients [9,10]. Multiple factors, including those of immunological, genetic, endocrine and environmental origin, contribute to the above condition. The phenomenon of epitope spreading can be used to explain this coexistence or this shift. Epitope spreading is a phenomenon in which immune responses can spread over the disease course to recognize epitopes that are different from the original target [11]. If it occurs in the same molecule, this is termed “intramolecular epitope spreading” [12,13]. When responses begin to target epitopes on other proteins, this is termed “intermolecular epitope spreading” [14,15].
The purpose of this study was to examine first the epitopes of anti-Dsg1 and anti-Dsg3 Abs from patients and healthy subjects in order to investigate whether there is any difference in Abs binding sites between groups and secondly to identify potential autoreactive HLA II-restricted helper T cell epitopes shared among Dsg1+/Dsg3+ subjects that could explain the simultaneous occurrence of these auto-Abs. Indeed, it has been reported that both qualitative and quantitative genetic variation of MHC class II molecules may influence susceptibility to endemic pemphigus foliaceus [16].
MATERIALS AND METHODS
Population’s demographics characteristics
PF patients were diagnosed on clinical, histological (acantholysis in the upper epidermis either in the granular layer or immediately below with subcorneal bullous formation) and immunological criteria (cell surface staining) of PF [18].
Serological analysis
Simultaneously, in both groups, antibodies to Dsg 1, Dsg 3 and BP180 (also known as BP antigen 2, the target antigen in bullous pemphigoid) were measured by an ELISA kits (MBL, Nagoya, Japan) for quantitative analysis as previously described [20].
According to the manufacturer’s instructions, index values >20 were considered positive for anti-Dsg1, anti-Dsg3 and anti-BP180.
Recombinant proteins and Immunoprecipitation-Immunoblotting analysis (IP-IB)
Immunoprecipitation was carried out by mixing a 20 ml bed volume of Protein G Sepharose 4 Fast Flow (GE Healthcare®, Uppsala, Sweden), 200 ml of culture supernatant for each protein, and 2 ml of each serum. In some experiments, the culture supernatant was treated with Ethylene Diamine Tetraacetic Acid (EDTA) at 5 mM for 1 h to chelete calcium. The mixture was incubated at 48°C overnight with gentle rotation and then washed five times with Tris-buffered saline containing 1 mM calcium chloride and 0.05% Tween 20. Immunoprecipitated proteins were then re-suspended in Sodium Dodecyl Sulfate (SDS) sample buffer, fractionated by SDS Polyacrylamide Gel Electrophoresis (SDS - PAGE), and labeled using a horseradish peroxidase-conjugated anti-E-tag Ab (GE Healthcare, Buckinghamshire, UK). Proteins were visualized using Western Lightning Chemiluminescence Reagent Plus (PerkinElmer LAS, Shelton, CT, USA) and by autoradiography.
HLA class II genes
HLA II genes encoded by the DRB1 and DQB1 loci were identified by Polymerase Chain Reaction (PCR) with Sequence-Specific Primers (PCR-SSP), as described previously [22].
RESULTS
Serological analysis (Table 1)
The ELISA for Dsg1 and Dsg3 was positive for all patients and healthy blood donors. Interestingly, the ELISA for BP180 was also positive in 2 healthy blood donors.
Epitope mapping analysis (Figure 1)
We also analyzed the epitope profile against Dsg3 autoantigen for all sera. Interestingly, IP-IB analysis of epitopes of all sera showed a profile skewed towards the C-terminal EC5 domain of Dsg3 (Figure 1b). It should be noted that the reaction of the healthy blood donors sera was weaker than the reaction of the PF patients sera.
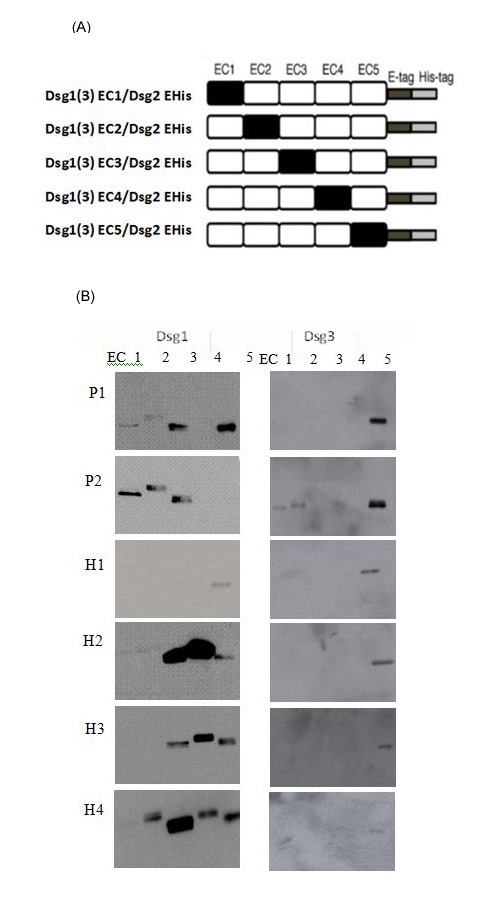
Immunogenetics analysis (Table 1)
According to previous study, HLA-DRB1*04 and DQB1*0302 are the risk alleles and HLA-DRB1*11/13 and DQB1*0301/06 are the protective alleles associated to endemic PF in Tunisia [22]. For PV disease, HLA-DRB1*04/08/14 and DQB1*0302 are statistically significant susceptibility factors where HLA-DRB1*03/07/13 and DRB1*15 may be protective alleles [23,24]. Similarly, genetic factors are also reported involved in the occurrence of BP; HLA-DRB1*04/11 and DQB1*03 alleles are suggestive to be two major BP susceptibility factors where HLA-DQB1*06 may be protective allele [7,25,26].
| Subjects | Demographics | Serological Studies | MHCII | |||||||
| IIF | ELISA | 1st Haplotype | 2nd Haplotype | |||||||
| Age | Sex | Dsg1 | Dsg3 | BP180 | DRB1 | DQB1 | DRB1 | DQB1 | ||
| Group I: Healthy blood donors | ||||||||||
| H1 | 26 | H | - | 30.5 | 22.1 | 43 | 13 | 02 | 03 | 06 |
| H2 | 48 | H | - | 36.3 | 22 | 27.5 | 13 | 06 | 13 | 06 |
| H3 | 80 | F | - | 81 | 25.1 | 5.7 | 11 | 0301 | 11 | 301 |
| H4 | 27 | F | - | 81.2 | 63 | 4.4 | 03 | 0301 | 14 | 04 |
| Group II: PF patients | ||||||||||
| P1 | 45 | F | - | 37.3 | 23.4 | 9 | 04 | 0302 | 14 | 05 |
| P2 | 30 | F | + | 164.5 | 33.7 | 6 | 04 | 0302 | 13 | 06 |
Table 1: Characterization of healthy blood donors and Pemphigus Foliaceus (PF) patients with antibodies to PF, Pemphigus Vulgaris (PV) and Bullous Pemphigoid (BP).
Group I: Healthy Blood Donors
Group II: PF Patients
DISCUSSION
Based on the specificity of the targeted antigens, several clinically and immune serologically distinct autoimmune bullous skin disorders have been characterized. Pemphigus and pemphigoid are an autoimmune blistering disorder with a complex etiology involving an interplay of genetic as well as environmental factors, most of which remain unknown. Pemphigus and pemphigoid are considered to be prototypic bullous disorders based on their well- defined immune response-mediated pathogenesis [27-29]. In PF, PV and BP, autoreactive CD4+T lymphocytes that are probably essential in initiating the autoimmune response recognize different epitopes of the extracellular portions of Dsg1, Dsg3 and BP180, respectively. Activation of autoreactive T cells in PF, PV and BP is restricted by distinct HLA class II alleles that are prevalent in individuals with these disorders. Autoreactive T cells are not only found in patients but have also been detected in healthy individuals [30]. In healthy individuals, a subset of autoreactive T cells with high regulatory function was also identified suggesting that the occurrence of autoimmune bullous disorders may be linked to a dysfunction of Tregs. Dsg3- and BP180-reactive T cells produce Th2 cytokines such as IL-4, IL-5, and IL-13 and facilitating immunoglobulin class switching from non-pathogenic (IgG1) to pathogenic (IgG4) autoantibodies, which are specially seen in active stages of these disorders.
Zakka et al., studied the relationship between target antigens and Major Histocompatibility Complex (MHC) class II genes in producing two pathogenic antibodies simultaneously [31]. Using computer models, they present the hypothesis that two autoantibodies are produced simultaneously due may be to the phenomena of epitope spreading through four possible scenarios. (i) One antigen molecule may contain at least two epitopes that have a certain point of similarity or overlapping sequence and can activate two different T cells and consequently stimulate two different B cells when presented by two different MHC II genes that may result in the simultaneous presence of two separate pathogenic autoantibodies. (ii) Antigen Presenting Cells (APC) may internalize two separate antigens, and then activate two different T cells that are specific to any one of these antigens. (iii) Two antigens may cluster in a single macromolecule complex, all of which are internalized by a single APC. This APC then activates two different T cells, making each specific for one subset of the macromolecule. The T cells would then solicit B cells and result in the production of two different autoantibodies. (vi) An autoimmune disease may cause tissue damage and inflammation in the surrounding tissue. This exposes the immune system to a previously sequestered epitope. This epitope becomes internalized by an APC, presented to a T cell that enlists B cells and produces a second antibody.
It has been also suggested that quantitative as well as qualitative variation of HLA class II molecules have an effect on the risk of an individual developing PF [16,32].
Flores et al., reported in 2012 that multiple, perhaps cross reactive, keratinocyte cadherins are recognized by Fogo Selvagem patients; the Brazilian endemic form of Pemphigus; and endemic controls. Authors suggested the relevance of the environment surrounding the patients and the endemic controls as a source of sensitizing antigen(s) that lead to the production of these autoantibodies through three possible explanations [9]. (i) The putative environmental antigen(s) may share cross-reactive epitopes with all the keratinocyte cadherins, thus triggering independent populations of autoantibodies with specificities to each of the keratinocyte cadherin. (ii) One or all of the environmentally-elicited anti-keratinocyte cadherin autoantibodies recognize crossreactive epitopes due to structural homologies among these keratinocyte cadherins. (iii) Finally, once an initial autoantibody response is triggered against one of these cadherins (primary response), this response may be followed by autoantibody responses against other keratinocyte cadherins by the epitope spreading mechanism.
This study demonstrates that the associations result from the effect of genetically controlled levels of candidate’s genes such as HLA II and production of autoantibodies are necessary for developing an autoimmune disease. The HLA DRB1* and DQB1* alleles influence susceptibility to pemphigus or pemphigoid and may contribute to severity and type of disease but cooperation of environmental, hormonal, and immunological factors and background are crucial to induce and activate any of these diseases.
REFERENCES
- Nousari HC, Anhalt GJ (1999) Pemphigus and bullous pemphigoid. Lancet 354: 667-672.
- Blank M, Gisondi P, Mimouni D, Peserico A, Piaserico S, et al. (2006) New insights into the autoantibody-mediated mechanisms of autoimmune bullous diseases and urticaria. Clin Exp Rheumatol 24: 20-25.
- Stanley JR, Amagai M (2006) Pemphigus, bullous impetigo, and the staphylococcal scalded-skin syndrome. N Engl J Med 355: 1800-1810.
- Schmidt E, Dähnrich C, Rosemann A, Probst C, Komorowski L, et al. (2010) Novel ELISA systems for antibodies to desmoglein 1 and 3: correlation of disease activity with serum autoantibody levels in individual pemphigus patients. Exp Dermatol 19: 458-463.
- Peterson JD, Chang AJ, Chan LS (2007) Clinical evidence of an intermolecular epitope spreading in a patient with pemphigus foliaceus converting into bullous pemphigoid. Arch Dermatol 143: 272-274.
- Fridkis-Hareli M (2008) Immunogenetic mechanisms for the coexistence of organ-specific and systemic autoimmune diseases. J Autoimmune Dis 5: 1.
- Zakka LR, Reche PA, Ahmed AR (2012) The molecular basis for the presence of two autoimmune diseases occurring simultaneously--preliminary observations based on computer analysis. Autoimmunity 45: 253-263.
- Arteaga LA, Prisayanh PS, Warren SJ, Liu Z, Diaz LA, et al. (2002) A subset of pemphigus foliaceus patients exhibits pathogenic autoantibodies against both desmoglein-1 and desmoglein-3. J Invest Dermatol 118: 806-811.
- Di Zenzo G, Zambruno G, Borradori L (2012) Endemic pemphigus foliaceus: towards understanding autoimmune mechanisms of disease development. J Invest Dermatol 132: 22499-22502.
- Flores G, Culton DA, Prisayanh P, Qaqish BF, James K, et al. (2012) IgG autoantibody response against keratinocyte cadherins in endemic pemphigus foliaceus (fogo selvagem). J Invest Dermatol 132: 2573-2580.
- Tchernev G, Orfanos CE (2006) Antigen mimicry, epitope spreading and the pathogenesis of pemphigus. Tissue Antigens 68: 280-286.
- Li N, Aoki V, Hans-Filho G, Rivitti EA, Diaz LA (2003) The role of intramolecular epitope spreading in the pathogenesis of endemic pemphigus foliaceus (fogo selvagem). J Exp Med 197: 1501-1510.
- Salato VK, Hacker-Foegen MK, Lazarova Z, Fairley JA, Lin MS, et al. (2005) Role of intramolecular epitope spreading in pemphigus vulgaris. Clin Immunol 116: 54-64.
- Pigozzi B, Peserico A, Schiesari L, Alaibac M (2008) Pemphigus foliaceus evolving into pemphigus vulgaris: a probable example of 'intermolecular epitope spreading' confirmed by enzyme-linked immunosorbent assay study. J Eur Acad Dermatol Venereol 22: 242-244.
- Recke A, Rose C, Schmidt E, Bröcker EB, Zillikens D, et al. (2009) Transition from pemphigus foliaceus to bullous pemphigoid: intermolecular B-cell epitope spreading without IgG subclass shifting. J Am Acad Dermatol 61: 333-336.
- Piovezan BZ, Petzl-Erler ML (2013) Both qualitative and quantitative genetic variation of MHC class II molecules may influence susceptibility to autoimmune diseases: the case of endemic pemphigus foliaceus. Hum Immunol 74: 1134-1140.
- Abida O, Kallel-Sellami M, Joly P, Ben Ayed M, Zitouni M, et al. (2009) Anti-desmoglein 1 antibodies in healthy related and unrelated subjects and patients with pemphigus foliaceus in endemic and non-endemic areas from Tunisia. J Eur Acad Dermatol Venereol 23: 1073-1078.
- Jarzabek-Chorzelska M, Jablonska S, Kolacinska-Strasz Z, Sulej I (2002) Immunopathological diagnosis of pemphigus foliaceus. Dermatology 205: 413-425.
- Toumi A, Saleh MA, Yamagami J, Abida O, Kallel M, et al. (2013) Autoimmune reactivity against precursor form of desmoglein 1 in healthy Tunisians in the area of endemic pemphigus foliaceus. J Dermatol Sci 70: 19-25.
- Ishii K, Amagai M, Hall RP, Hashimoto T, Takayanagi A, et al. (1997) Characterization of autoantibodies in pemphigus using antigen-specific enzyme- linked immunosorbent assays with baculovirus-expressed recombinant desmogleins. J Immunol 159: 2010-2017.
- Chan PT, Ohyama B, Nishifuji K, Yoshida K, Ishii K, et al. (2010) Immune response towards the amino-terminus of desmoglein 1 prevails across different activity stages in nonendemic pemphigus foliaceus. Br J Dermatol 162: 1242-1250.
- Svecova D, Parnicka Z, Pastyrikova L, Urbancek S, Luha J, et al. (2015) HLA DRB1* and DQB1* alleles are associated with disease severity in patients with pemphigus vulgaris. Int J Dermatol 54: 168-173.
- Párnická Z, Švecová D, Javor J, Shawkatová I, Buc M, et al. (2013) High susceptibility to pemphigus vulgaris due to HLA-DRB1*14:54 in the Slovak population. Int J Immunogenet 40: 471-475.
- Oyama N, Setterfield JF, Powell AM, Sakuma-Oyama Y, Albert S, et al. (2006) Bullous pemphigoid antigen II (BP180) and its soluble extracellular domains are major autoantigens in mucous membrane pemphigoid: the pathogenic relevance to HLA class II alleles and disease severity. Br J Dermatol 154: 90-98.
- Gao XH, Winsey S, Li G, Barnardo M, Zhu XJ, et al. (2002) HLA-DR and DQ polymorphisms in bullous pemphigoid from northern China. Clin Exp Dermatol 27: 319-321.
- Hertl M (2000) Humoral and cellular autoimmunity in autoimmune bullous skin disorders. Int Arch Allergy Immunol 122: 91-100.
- Stanley JR (1989) Pemphigus and pemphigoid as paradigms of organ-specific, autoantibody-mediated diseases. J Clin Invest 83: 1443-1448.
- Borradori L, Sonnenberg A (1996) Hemidesmosomes: roles in adhesion, signaling and human diseases. Curr Opin Cell Biol 8: 647-656.
- Hertl M, Eming R, Veldman C (2006) T cell control in autoimmune bullous skin disorders. J Clin Invest 116: 1159-1166.
- Zakka LR, Keskin DB, Reche P, Ahmed AR (2010) Relationship between target antigens and Major Histocompatibility Complex (MHC) class II genes in producing two pathogenic antibodies simultaneously. Clin Exp Immunol 162: 224-236.
- Paisansinsup T, Deshmukh US, Chowdhary VR, Luthra HS, Fu SM, et al. (2002) HLA class II influences the immune response and antibody diversification to Ro60/Sjogren’s syndrome-A: heightened antibody responses and epitope spreading in mice expressing HLA-DR molecules. J Immunol 168: 5876-5884.
Citation: Toumi A, Abida O, Masmoudi H (2017) Antibodies against Desmoglein 1, Desmoglein 3 and BP180 in Healthy Tunisians Blood Donors without Immunopathological Phenotype. J Clin Dermatol Ther 4: 024.
Copyright: © 2017 Hatem Masmoudi, et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.