Plasma Citrate Homeostasis: How It Is Regulated; And Its Physiological and Clinical Implications. An Important, But Neglected, Relationship in Medicine
*Corresponding Author(s):
Leslie C CostelloThe Department Of Oncology And Diagnostic Sciences, School Of Dentistry, University Of Maryland, Baltimore, MD, 21201, United States
Tel:+1 4107067618,
Email:lcostello@umaryland.edu
Abstract
The homeostatic maintenance of a normal plasma citrate concentration is an important factor in humans and in animals; and is required for many normal physiological activities. Dysregulation of normal plasma citrate presents pathophysiological hypocitricemic or hypercitricemic conditions. This can lead to clinical consequences in many areas of medicine; such as impaired blood clotting, altered acid/base status, impaired neuromuscular/cardiac activities, hypocitraturia and stone formation, bone disorders with loss of bone strength and increased fractures, hypocitricemia of surgical stress. These important implications of citrate relationships have been largely ignored by the contemporary clinical and biomedical community; to the extent that it is not even described in most current textbooks and review papers. This review describes the physiological, endocrine, and metabolic relationships in the normal regulation and maintenance of plasma citrate; and presents some important clinical consequences of its dysfunctional maintenance. The importance of bone, kidney and liver activities in the maintenance of normal plasma citrate is described along with the citricemic roles of parathyroid hormone, calcitonin and vitamin D. These factors and relationships are presented as the contemporary understanding of the integrated regulation of plasma citrate as the basis for its clinical importance in medicine. The exclusion of these citrate relationships leads to misunderstanding and misrepresentation of physiological and clinical conditions in many issues in medicine and paramedicine areas. The intent of this review is to revive the interest and support for research to address the many unknown and speculative issues of plasma citrate regulation and its important clinical implications. This is in the best interest of the medical community and the public-at-large.
Keywords
INTRODUCTION
Plasma citrate is maintained at a normal constant concentration in humans and animals. In humans the normal plasma citrate concentration is within a range of ~100-150µM. Exposure to conditions that tend to increase or decrease the plasma citrate beyond this normal range will trigger physiological, endocrinological, and metabolic responses that restore and maintain the normal plasma citrate concentration. If the plasma citrate concentration is sustained at a decreased or an increased concentration beyond the normal range, a hypocitricemia or hypercitricemia exists which is indicative of a pathophysiological status. Such conditions can impose clinical consequences (described below). The maintenance and regulation of plasma citrate and its physiological/pathophysiological/clinical implications remain largely unknown, speculative, and/or overlooked by most of the contemporary clinical and biomedical research community.
The intent of this review is to bring attention to the importance of normal citrate homeostasis. A background is presented of the existing information regarding the factors and mechanisms involved in the regulation and maintenance of citrate homeostasis. The pathophysiological and clinical consequences of dysregulated plasma citrate are described. As best that we can determine, this is the first review that describes and integrates these citrate relationships collectively in the context of the homeostasis of plasma citrate and its implications in medicine. Hopefully, this will stimulate interest and support for research, which are necessary to establish and understand the important role of citrate in the normal functional activities of the body; and the clinical consequences of disrupted citrate homeostasis.
THE NORMAL SOURCE AND REMOVAL OF PLASMA CITRATE
It is evident that there exists a balance between citrate that enters circulation and citrate that is removed from circulation, which results in the maintenance of the plasma citrate concentration. The prevailing views over time have included dietary citrate as the exogenous source, bone as the reservoir source of citrate, and tissue/cellular metabolism as an endogenous de novo source of plasma citrate. The removal of citrate from circulation has been considered to result from its urinary excretion, from its deposition into bone, and from its uptake and metabolism by various cells/tissues. However, much of the integrated role of these factors in the maintenance of plasma citrate has remained speculative and also inaccurate. The following provides a description of each of the factors as represented in figure 1, which leads to our current understanding of citrate homeostasis.
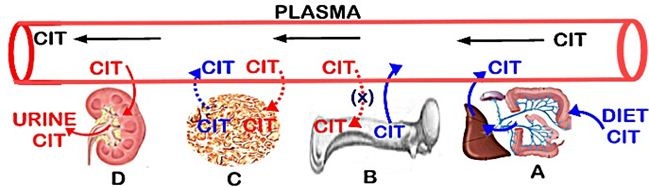
THE MECHANISMS OF THE ROLE OF BONE AND KIDNEYS IN THE MAINTENANCE OF PLASMA CITRATE
Bone as the major source of citrate in plasma
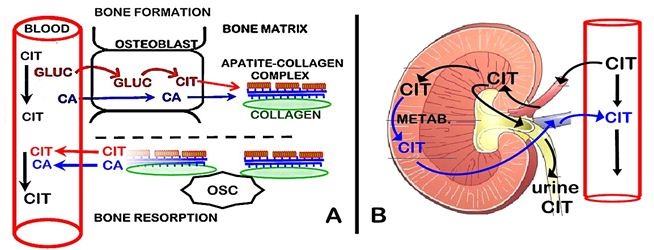
During bone resorption, both citrate and calcium are released from bone into plasma. This provides a de novo source of citrate for the maintenance of the plasma concentration; whereas the calcium that enters the plasma is calcium that is originally derived from plasma (i.e., calcium turnover). Because bone turnover (bone formation↔bone resorption) occurs throughout life, it provides a continual source of new citrate as a major factor in plasma homeostasis. This is a new understanding of the important role of the osteoblasts and bone turnover in citrate homeostasis, which has major implications regarding the hormonal and other factors that regulate citrate homeostasis, and its clinical implications.
The renal clearance of citrate
Since normal renal function involves some level of obligatory citrate clearance (~500 mg/day), that loss of plasma citrate must be compensated by an equal addition of citrate into plasma. Dietary citrate, when available contributes to the replacement of citrate. However, bone resorption provides the citrate that is required to achieve the normal plasma concentration of citrate; and osteoblast de novo production of citrate refurbishes bone citrate during bone formation.
THE HORMONAL REGULATION OF PLASMA CITRATE CONCENTRATION
Exogenous and endogenous conditions and agents continually impose changes in the plasma citrate concentration. Such changes must be countered by adjustments in the citrate that enters the plasma and/or the citrate that is removed from the plasma to maintain the normal plasma citrate concentration. This requires endocrine monitoring of the plasma citrate concentration, and its hormonal response to maintain the normal plasma concentration; i.e., “citricemic” hormones. Unfortunately, aside from the early studies over the period of ~1950-1980, the focus on the physiological and endocrinological regulation of citrate homeostasis has been largely ignored. So, we must rely predominantly on the information provided in the early reports and integrated with the limited more recent reports as the basis for the relevant contemporary issues.
Parathyroid Hormone (PTH) and Calcitonin (CT); the major citricemic hormones
Parathyroid hormone:
PTH effects on bone citrate:
All of these hypocalcemia effects are accompanied by the corresponding CT hypocitricemia effects. Studies during 1970-1973 by Komarkova et al., [26] and Costello et al., [27,28] established the CT which induces hypocitricemia along with hypocalcemia. This was corroborated by Natelson et al in 1979 [29], who reported that administration of calcitonin in rabbits results in hypocitricemia. Since then, no other reports exist (as best we could determine) of the hypocitricemic effect of CT. Thus, a poor understanding of the mechanisms of CT regulation of plasma citrate and its pathophysiological implications currently exists. However, it is reasonably established that the citricemic effects are achieved by CT actions on bone and on renal citrate clearance, as are the calcemic effects.
The new understanding (Figure 3A) of osteoblast citrate production during bone formation and its independence of the calcium source, negates the CaCit coupling of the action of CT. Since CT exhibits direct effects on the osteoblasts during osteogenesis [30-34], it becomes extremely likely that CT promotes osteoblast production of citrate along with its mineralization during bone formation. Since no reported studies exist regarding CT effects on osteoblast citrate production, this is an essential issue that needs to be established.
The new understanding (Figure 3A) of osteoblast citrate production during bone formation and its independence of the calcium source, negates the CaCit coupling of the action of CT. Since CT exhibits direct effects on the osteoblasts during osteogenesis [30-34], it becomes extremely likely that CT promotes osteoblast production of citrate along with its mineralization during bone formation. Since no reported studies exist regarding CT effects on osteoblast citrate production, this is an essential issue that needs to be established.
It is notable that despite the focus on CT hypocalcemia, the effect of CT in the renal clearance of calcium remains conflicted. Some reports [35] conclude that CT results in increased renal clearance of calcium; other reports [36] conclude that CT results in decreased renal clearance of calcium. Since the latter is inconsistent with its being a factor in the hypocalcemic role of CT, it is presumed that the effect of CT on bone is the major action for CT hypocalcemia. Nevertheless, the renal handling of citrate is independent of calcium and a coupled clearance of calcium with citrate.
There are conditions in which plasma citrate regulation exists independent of corresponding plasma calcium changes. For example, vitamin D promotes hypercitricemia, which involves bone and renal effects; whereas no corresponding effect on plasma calcium (and other parameters) occurs (described below). It is also notable that the administration of high levels of citrate results in a transient increase in plasma citrate and increased citraturia followed by a normocitricemia; without involvement of an accompanying calcemic response.
In our studies in humans and rats [38,39], we observed that the surgical stress hypocitricemic response did not involve any increase in the renal clearance of citrate; and it did not involve either PTH or CT. Consequently, we pursued the possibility of the liver clearance of plasma citrate being involved in the surgical stress hypocitricemia in rats. For this, we determined the hepatic Citrate Extraction Coefficient (CEC). The liver receives its blood supply from the Hepatic Artery (HA) and the Hepatic Portal Vein (HPV). In humans, HA provides ~25% and HPV~75% of the liver blood supply. In rats, the values are ~33% and 67%, respectively. Therefore the CEC equation for rats requires the calculation of the citrate “Arterial” concentration = (0.33HA) + (0.67HPV); then
CEC = (“Arterial” conc + Hepatic Vein conc) / “Arterial” conc”
These conditions (Figure 4) demonstrate that the liver citrate extraction coefficient is negligible in the normal animals; whereas surgical animals exhibit a CEC~0.55. Unlike the earlier reports, this study employs in situ liver citrate extraction from circulation which establishes the minimal, if any, normal hepatic citrate extraction. Thus, surgical stress induces a major hepatic citrate clearance of 55% of the citrate in the plasma that it receives. In the absence of increased renal clearance of citrate, the increased hepatic clearance of citrate is the major factor that results in surgical stress hypocitricemia. This becomes evident when comparing this hepatic citrate clearance to renal citrate clearance which is generally considered to be the major factor in the removal of citrate from circulation. Our rat studies showed that the normal renal citrate extraction is ~25% as compared to the liver citrate extraction of ~55% following surgery. The relative blood flow for the kidneys versus liver is ~1:1.4. Therefore liver citrate clearance following surgery is ~3-fold greater than normal renal citrate clearance. It also becomes evident that this high hepatic citrate clearance provides an explanation for surgical hypocitricemia over-riding the hypercitricemic effects of excessive parathyroid hormone and the hypocitricemic effects of calcitonin. This is an important liver relationship that had never been recognized.
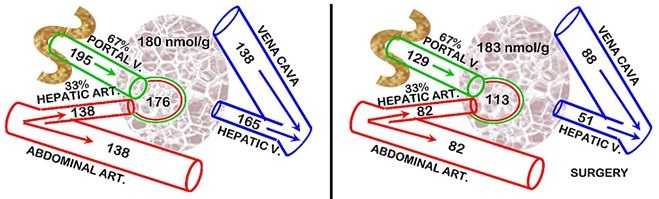
It is apparent from this study that the liver, under normal conditions is not significantly involved in the clearance of plasma citrate for the homeostatic maintenance of plasma citrate concentration. However, under specific conditions the hepatic clearance of citrate and the activities of the hepatic cells can be a major factor in maintaining the normal plasma citrate concentration.
THE PATHOPHYSIOLOGICAL AND CLINICAL CONSEQUENCES OF DISRUPTED PLASMA CITRATE HOMEOSTASIS
The plasma citrate-plasma calcium connection
An important example is the clinical consequences of the use of citrated blood for massive blood transfusion and blood exchange in adults and children. During the period of ~1945-1965, the introduction of massive blood transfusion as a medical procedure resulted in the identification of the clinical entity characterized as “citric acid (citrate) intoxication” [43-45]. The condition is a severe hypercitricemia (plasma citrate concentration is increased by ~10-fold or greater); which decreases the Ca++ by ~50% or more. The consequences included impaired blood coagulation; extended bleeding time, impaired cardiac function and EKG, hypotension; tetany; metabolic alkalosis, and in some cases, death. Since then, the issue of “citric acid intoxication”, as indicated by the sharp decline in published reports, had not received much attention. The recent increasing applications of massive blood transfusion for organ transplants, major surgical tissue resections, and for blood exchange has renewed the interest in the issue of citric acid intoxication, as is evident from recent reports such as [46-48]. The potential for citric acid intoxication is reflected in the conclusion [44] that “it is possible to predict with assurance that citric acid intoxication is likely to occur during multiple transfusions of citrated blood in patients during extremely rapid and prolonged infusions of citrated blood or plasma”. The implications of citrated blood for pediatric as well as adult conditions warrant its consideration and management.
The plasma citrate; renal clearance of citrate; calcium stone formation
Conditions that decrease the concentration of citrate in the tubular fluid will lead to hypocitraturia and stone formation. This includes hypocitricemia in which the concentration of citrate entering the glomerular filtrate limits its availability in urine formation. Hypocitricemia due to insufficient dietary source of citrate can be such a factor.
However, hypocitraturia often occurs in the presence of normocitricemia and hypercitricemia. This is represented by acid–base status, which is the most important determinant of urinary citrate excretion. Metabolic and cellular acidosis is a major cause of hypocitraturia. Acidosis increases the transport of citrate from the tubular fluid into the tubular cells, and it increases the cellular metabolic utilization of the citrate. Consequently, urinary citrate is markedly decreased. Conditions that lead to metabolic acidosis or cellular acidosis (e.g., potassium deficiency, excess sodium, ketosis, exercise) will promote hypocitraturia.
A different effect is exemplified by hyperparathyroidism as a cause of hypercalcemia along with hypocitraturia and stone formation. In this case, the renal clearance of citrate is decreased; and the reabsorption of citrate from the tubular fluid back into circulation is increased. This differs from acidotic hypocitraturia, in which the reabsorption of citrate back into circulation is decreased. This difference implies that tubular cell uptake of citrate occurs in both conditions, but the cellular utilization of citrate is inhibited in response to parathyroid hormone (4.1.1b above).
The complexity of interacting factors in the renal clearance of citrate mandates the need for continued research into these issues.
Citrate dysregulation and bone disorders
VITAMIN D AND THE MAINTENANCE OF PLASMA CITRATE CONCENTRATION
Although not a hormonal factor that directly responds to the plasma citrate concentration, there are numerous reports [51-56] that describe the role of vitamin D in the maintenance of the normal plasma concentration. Vitamin D deficiency results in hypocitricemia and administration of vitamin D increases the plasma citrate concentration. Along with the increase in plasma citrate, vitamin D also increases kidney tissue and bone citrate concentrations. The citrate effects of vitamin D are not dependent or associated with calcium or phosphate status, alkalosis, or other such conditions; thereby demonstrating vitamin D as a specific “hypercitricemic” agent.
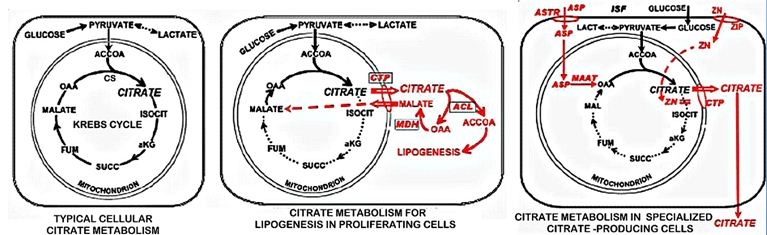
These relationships are clinically implicated in vitamin D-deficient rickets, which results from the insufficient plasma citrate availability, particularly in infants and juveniles [1,52,57,58]. The rachitic bone is fragile and highly susceptible to fracture. The treatment with supplemental citrate and vitamin D restores the bone strength and resistance to fracture. Despite this important relationship, the mechanism by which vitamin D manifests its increased tissue citrate effect is unknown. Vitamin D induced increase in citrate production results from the inhibition of citrate oxidation, and is mimicked by fluoroacetate inhibition of citrate oxidation [56,59,60]. Fluoroacetate is a specific inhibitor of m-aconitase activity, which then prevents citrate oxidation via the Krebs cycle. Cells that conserve rather than oxidize citrate, exhibit high zinc levels, which specifically inhibits m-aconitase activity [14], and this applies to osteoblast citrate production during bone formation (Figure 2) [3,4]. It is plausible to expect that the vitamin D citrate-producing effect might involve this mechanism. Because of its important implications, the mechanism and factors involved in vitamin D induced citrate production need to be established.
SUMMARY
• Citrate homeostasis is essential for manifestation of normal physiological activities; and its dysregulation has pathophysiological implications and clinical consequences
• The normal plasma concentration is achieved mainly by bone turnover which provides the major source of plasma citrate, and renal citrate clearance for the removal of citrate; the combination of which maintains normocitricemia
• The above is regulated by endocrine citricemic hormones in response to the plasma citrate concentration, either in combination with, or independent of, calcium regulation. Parathyroid hormone and calcitonin are respectively hypercitricemic and hypocitricemic hormones, which regulate the bone and renal activities to maintain normocitricemia
• Vitamin D is a major specific hypercitricemic agent which is achieved by promoting the activities of bone and kidney in producing and conserving citrate
• The liver does not normally remove citrate from plasma; but under specific conditions increased hepatic clearance of citrate is a major factor
• The dysregulation of plasma citrate homeostasis has pathophysiological/and clinical consequences which include: plasma calcium related defective blood clotting and impaired neuromuscular/cardiac activity: hypocitraturia and renal stone formation; bone disorders characterized by bone fragility and fractures (such as juvenile rickets and osteoporosis)
Unfortunately, the normal and dysfunctional implications of citrate homeostasis have been largely ignored by the contemporary clinical and biomedical community. The intent of this review is to bring attention to the importance of citrate homeostasis in medicine, and the need for enhanced research and funding to address the many unknown and unrecognized issues and relationships. It is in the best interest of the medical community and the public-at large that this should no longer remain a neglected area of interest and research.
ACKNOWLEDGEMENT
The studies of LCC and RBF presented in this review were supported by NIH grants AR064808, DK076783 and CA79903.
REFERENCES
- Dickens F (1941) The citric acid content of animal tissues, with reference to its occurrence in bone and tumour. Biochem J 35: 1011-1023.
- Costello LC, Chellaiah M, Zou J, Franklin RB, Reynolds MA (2014) The status of citrate in the hydroxyapatite/collagen complex of bone; and its role in bone formation. J Regen Med Tissue Eng 3: 4.
- Franklin RB, Chellaiah M, Zou J, Reynolds MA, Costello LC (2014) Evidence that Osteoblasts are Specialized Citrate-producing Cells that Provide the Citrate for Incorporation into the Structure of Bone. Open Bone J 6: 1-7.
- Costello LC, Franklin RB, Reynolds MA, Chellaiah M (2012) The Important Role of Osteoblasts and Citrate Production in Bone Formation: “Osteoblast Citration” as a New Concept for an Old Relationship. Open Bone J: 4.
- Costello LC, Chellaiah MA, Zou J, Reynolds MA, Franklin RB (2015) In vitro BMP2 stimulation of osteoblast citrate production in concert with mineralized bone nodule formation. J Regen Med Tissue Eng 4.
- Zuckerman JM, Assimos DG (2009) Hypocitraturia: Pathophysiology and Medical Management. Rev Urol 11: 134-144.
- Simpson DP (1983) Citrate excretion: a window on renal metabolism. Am J Physiol 244: 223-234.
- Zacchia M, Preisig P (2010) Low urinary citrate: an overview. J Nephrol 16: 49-56.
- Hamm LL (1990) Renal handling of citrate. Kidney Int 38: 728-735.
- Wong GL, Kent GN, Ku KY, Cohn DV (1978) The interaction of parathormone and calcium on the hormone-regulated synthesis of hyaluronic acid and citrate decarboxylation in isolated bone cells. Endocrinology 103: 2274-2282.
- Luben RA, Cohn DV (1976) Effects of parathormone and calcitonin on citrate and hyaluronate metabolism in cultured bone. Endocrinol 98: 413-419.
- Wong GL (1983) Actions of parathyroid hormone and 1,25-dihydroxycholecalciferol on citrate decarboxylation in osteoblast-like bone cells differ in calcium requirement and in sensitivity to trifluoperazine. Calcif Tissue Int 35: 426-431.
- Wolinsky I, Cohn DV (1969) Oxygen uptake and 14CO2 production from citrate and isocitrate by control and parathyroid hormone-treated bone maintained in tissue culture. Endocrinol 84: 28-35.
- Costello LC, Liu Y, Franklin RB, Kennedy MC (1997) Zinc inhibition of mitochondrial aconitase and its importance in citrate metabolism of prostate epithelial cells. J Biol Chem 272: 28875-28881.
- Nagata N, Rasmussen H (1968) Parathyroid hormone and renal cell metabolism. Biochemistry 7: 3728-3733.
- Cohn DV, Smaich AF, Levy R (1966) The inhibition of respiration and phosphorylation in kidney mitochondria by parathyroid hormone administered in vivo. J Biol Chem 241: 889-894.
- Costello LC, Wynn R, Goldberg M (1966) In vitro effect of parathyroid extract on isocitrate oxidation by rat kidney mitochondria. Endocrinology 78: 614-620.
- Costello, LC, Darago LL (1964) Parathyroid extract inhibition of isocitrate oxidation by kidney mitochondria. Arch Biochem Biophys 108: 574-576.
- Costello, LC, Darago LL (1964) The in vivo effects of parathyroid extract on the oxidation of citrate and isocitrate by kidney mitochondria. Gen Comp Endocrinol 4: 159-163.
- Alvarez-Arroyo MV, Traba ML, Rapado A, de la Piedra C (1992) Role of citric acid in primary hyperparathyroidism with renal lithiasis. Urol Res 20: 88-90.
- Copp DH (1994) Calcitonin: discovery, development, and clinical application. Clin Invest Med 17: 268-277.
- Hirsch PF (1971) Thyrocalcitonin and its role in calcium regulation in mammals. J Exp Zool 178: 139-149.
- Copp DH (1969) Endocrine control of calcium homeostasis. J Endocrinol 43: 137-161.
- Zaidi M, Inzerillo AM, Moonga BS, Bevis PJ, Huang CL (2002) Forty years of calcitonin--where are we now? A tribute to the work of Iain Macintyre, FRS. Bone 30: 655-663.
- Kleeman CR, Massry SG, Coburn JW (1971) The clinical physiology of calcium homeostasis, parathyroid hormone, and calcitonin. I. Calif Med 114: 16-43.
- Komárková A, Bilyk I, Záhor Z, Czabanová V (1970) Thyrocalctionin and citric acid. Experientia 26: 196-198.
- Franklin R, Costello LC, Stacey R, Stephens R (1973) Calcitonin effects on plasma and urinary citrate levels in rats. Am J Physiol 225: 1178-1180.
- Costello LC, Stacey R, Stevens R (1971) Hypocitricemic effects of calcitonin, parathyroidectomy and surgical stress. Horm Metab Res 3: 120-125.
- Natelson S, Miletich DJ, Seals CF, Visintine DJ, Albrecht RF (1979) Clinical biochemistry of epilepsy. II. Observations on two types of epileptiform convulsions induced in rabbits with corticotropin. Clin Chem 25: 898-913.
- Farley JR, Wergedal JE, Hall SL, Herring S, Tarbaux NM (1991) Calcitonin has direct effects on 3[H]-thymidine incorporation and alkaline phosphatase activity in human osteoblast-line cells. Calcif Tissue Int 48: 297-301.
- Villa I, Dal Fiume C, Maestroni A, Rubinacci A, Ravasi F, et al. (2003) Human osteoblast-like cell proliferation induced by calcitonin-related peptides involves PKC activity. Am J Physiol Endocrinol Metab 284: 627-633.
- Wong GL, Cohn DV (1975) Target cells in bone for parathormone and calcitonin are different: enrichment for each cell type by sequential digestion of mouse calvaria and selective adhesion to polymeric surfaces. Proc Natl Acad Sci USA 72: 3167-3171.
- Farley JR, Tarbaux NM, Hall SL, Linkhart TA, Baylink DJ (1988) The anti-bone-resorptive agent calcitonin also acts in vitro to directly increase bone formation and bone cell proliferation. Endocrinology 123: 159-167.
- Farley JR, Dimai HP, Stilt-Coffing B, Farley P, Pham T, et al. (2000) Calcitonin Increases the Concentration of Insulin-Like Growth Factors in Serum-Free Cultures of Human Osteoblast-Line Cells. Calcified Tissue International 67: 247-254.
- Cochran M, Peacock M, Sachs G, Nordin (1970) Renal effects of calcitonin. Br Med J 17: 135-137.
- Carney S, Thompson L (1981) Acute effect of calcitonin on rat renal electrolyte transport. Am J Physiol 240: 12-16.
- Martin GR, Firschbein HE, Mulrayanan BJ, Neuman WF (1958) Concerning the Mechanism of Action of Parathyroid Hormone. II. Metabolic Effects1,2. J Am Chem Soc 80: 6201-6204.
- Costello LC, Balkissoon B, Stacey R (1973) Hypocitricemic effect of surgical stress. J Surg Res 15: 182-188.
- Costello LC, Fadika G, Stacey R (1977) Hepatic citrate changes in surgical stress. J Surg Res 22: 92-96.
- Vang OJ, Zemel R, Davidson M, Drapanas T (1966) Citrate metabolism by the isolated perfused liver. Arch Surg 93: 142-147.
- Martensson J (1940) On the citric acid metabolism in mammals. Acta Physiologica Scandinavica 1: 1-96.
- Sjostrom P (1937) Citric acid in the blood serum in the diagnosis of the diseases of the liver and bile ducts; a methodological, experimental, and clinical study. Acta Chir Scandinav 49.
- Wexler IB, Pincus JB, Natelson S, Lugovoy JK (1949) III. The fate of citrate in erythroblastotic infants treated with exchange transfusion1,2. J Clin Invest 28: 474-81.
- Bunker JP, Stetson JB, Coe RC, Grillo HC, Murphy AJ (1955) Citric acid intoxication. JAMA 157: 1361-1367.
- Yendt ER (1957) Citrate intoxication. Can Med Assoc J 76: 141-144.
- Ikeda T, Usui A, Matsumura T, Aramaki T, Hosoya T, et al. (2015) An autopsy case of death due to metabolic acidosis after citric acid ingestion. Leg Med (Tokyo) 17: 532-534.
- Li K, Xu Y (2015) Citrate metabolism in blood transfusions and its relationship due to metabolic alkalosis and respiratory acidosis. Int J Clin Exp Med 8: 6578-6584.
- B?çakç? Z, Olcay L (2014) Citrate metabolism and its complications in non-massive blood transfusions: association with decompensated metabolic alkalosis+respiratory acidosis and serum electrolyte levels. Transfus Apher Sci 50: 418-426.
- Hu YY, Rawal A, Schmidt-Rohr K (2010) Strongly bound citrate stabilizes the apatite nanocrystals in bone. Proc Natl Acad Sci USA 107: 22425-22429.
- Davies E, Müller KH, Wong WC, Pickard CJ, Reid DG, et al. (2014) Citrate bridges between mineral platelets in bone. Proc Natl Acad Sci USA 14: 1354-1363.
- Caudarella R, Fabio V, Angela B, Sergio S (2003) Citrate and mineral metabolism: Kidney stones and bone disease. Frontiers Biosci 8: 1084-1106.
- Nicolaysen R, Eeg-Larsen N (1953) The biochemistry and physiology of vitamin D. Vitamins & Hormones 11: 29-60.
- Nicolaysen R, Nordb R (1943) Calcium metabolism and citric acid. Acta Physiologica 5: 212-214.
- Freeman S, Chang TS (1950) Effect of thyroparathyroidectomy and vitamin D upon serum calcium and citric acid of normal and nephrectomized dogs. Am J Physiol 160: 341-347.
- Carlsson A, Hollunger G (1954) The effect of vitamin D on the citric acid metabolism. Acta Physiol Scand 31: 317-333.
- Steenbock H, Bellin SA (1953) Vitamin D and tissue citrate. J Biol Chem 205: 985-991.
- Harrison HE, Harrison HC (1952) Vitamin D and Citrate Metabolism: Studies on Rachitic Infants. Yale J Biol Med 24: 273-283.
- Harrison HC, Harrison HE, Park EA (1958) Vitamin D and citrate metabolism; effect of vitamin D in rats fed diets adequate in both calcium and phosphorus. Am J Physiol 192: 432-436.
- De Luca HF, Steenbock H (1957) An in vitro effect of vitamin D on citrate oxidation by kidney mitochondria. Science 126: 258.
- Potter VR, Busch H (1950) Citric acid content of normal and tumor tissues in vivo following injection of fluoroacetate. Cancer Res 10: 353-356.
Citation: Costello LC, Franklin RB (2016) Plasma Citrate Homeostasis: How It Is Regulated; And Its Physiological and Clinical Implications. An Important, But Neglected, Relationship in Medicine. J Hum Endocrinol 1: 005.
Copyright: © 2016 Leslie C Costello, et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.