Some Reflections on the Beginning of the Oldest Corals Rugosa
*Corresponding Author(s):
Yuri LatypovNational Scientific Center Of Marine Biology, Vladivostok, Russian Federation
Tel:+7 9136956205,
Email:yuri.latypov@mail.ru
Abstract
Provides a conceptual theory of origin Rugosa. Discusses the possibility of the oldest single corals from the Cambrian forebears. With specific examples showing ways of their development and transmission of hereditary traits. Discusses the validity of the terms “Tetracorallia” and “Rugosa” and their bilaterally.
Keywords
MATERIALS AND METHODS
CONCEPTUAL INTRODUCTION IN THE THEORY OF THE ORIGIN OF RUGOSA
First rugosa had only simple septa and full bottom with single additional plates, but by the beginning of the Late Ordovician plate tabulae acquire the ability to very wide variation, and by the end of the Late Ordovician kaliko blastes basal surface polyp’s single rugosa acquired the ability to secrete peripheral convex skeletal elements - septum (Paliphyllum). By the beginning of the Late Ordovician all types of microstructures - lamellar, fibrous, monakant, golokant, rabdakant, dimorfakant participated in the construction of the tabulae, septa, plate and akanthin septa. The ability to secrete lamellar sclerenchyma vertical and horizontal skeletal elements like thickening until their mutual fusion, was developed from the beginning rugosa stories and observed in many genera, or lesser greater extent until the end of their existence. A stereoplasmic growth layer (sclerenchyma cover) expanded also towards the remaining skeletal elements [1,5,6]. All such secondary thickenings were originally composed of crystalline fibrils and trabecular oriented parallel to the surface of a given skeletal element.
We can only speculate whether the evolution of life forms coral influenced change streptelasmatid internal structure or vice versa deliverance from primitive massive improved internal structure of the skeletal elements and development caused the growth of the species diversity of these rugosa that we find at the end of the single coral Late Llandovery. Even with the experimental data confirming this account with what some modern similar corals, we would not be able to transfer these results to streptelasmatid, since the nature of the appearance of single scleractinian metasept have in principle different from the method of laying new septum in single-band single fossil corals. But those elements evolution streptelasma tid that we can observe on the skeletal remains of these concrete rugosa remain undoubted facts allowing them uniquely interpret any professionals involved in the study of corals.
One way or another, but by the end of the Middle - Late Ordovician top single rugosa had an overwhelming number of morphological traits, vertical and horizontal skeletal elements, the microstructure of the entire arsenal of features that have occurred in the history of existence of the Middle Ordovician to the end of the Permian. Basic life forms were developed - a conical, cylindrical-conical, trochoid, turbidity. Vertical single skeletal elements deposited rugosa as plate and spiny, large and small septum. Partitions themselves since the beginning of the emergence possessed a high degree of variability in shape, curvature, length and width, as well as spatial orientation in the axial zone corallites. Various types of axial structures can form in the cup coral subtle and very prominent uplift. Oriented septum bilaterally pinnately or subradial released with size major protosepta that could pitch in fossula recess.
Along with this happening deliverance from sclerenchymal thickening of the skeletal elements, the development of septal apparatus of thick rare septum in numerous thin septa. Increasing the number and size interseptal cameras, converting the axial structure of the solid with central sclerenchyma raising through fungous- porous structure of the entanglements septa and tabulae in the axial wave septum, and then the release of the central area of any coral formations axis. Transformation complete rare tabulae in numerous incomplete convex interseptal lamellas, sometimes to proto dissepiments (Figure 1). This single form of existence and throughout the history of bilateral streptelasmatid orientation corallites their bodies and that at the time of their greatest prosperity become pronounced.
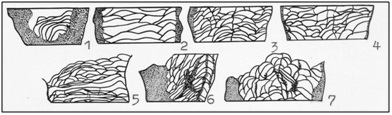 Figure 1: Improvement and specialization tabulae of Streptelasmatidae. 1-Kenophyllum, middle Ordovician, 2-Streptelasma, late Ordovician 3-Dinophyllum, Llandoveri, 4-Porfirieviella, late Llandoveri, 5-Pseudophaulactis, late Llandoveri, 6-Streptelasma, Llandoveri 7-Grewingkia, late Ordovician [1].
Figure 1: Improvement and specialization tabulae of Streptelasmatidae. 1-Kenophyllum, middle Ordovician, 2-Streptelasma, late Ordovician 3-Dinophyllum, Llandoveri, 4-Porfirieviella, late Llandoveri, 5-Pseudophaulactis, late Llandoveri, 6-Streptelasma, Llandoveri 7-Grewingkia, late Ordovician [1].From this perspective, it seems to me likely following scheme of single-band single rugosa (Figure 2).
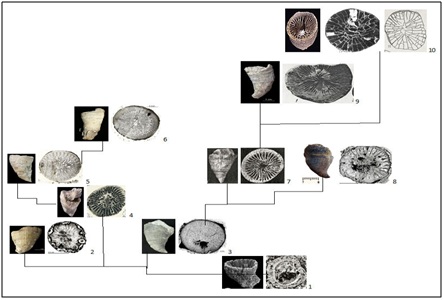 Figure 2: Scheme development of Streptelasmatidae. The appearance of the corallites, nature of its sclerenchymal deposits, septum and interseptal camera: 1-Cothonion, 485,5 Ma, 2-Primitophyllum, 3-Kenophyllum, 4-Dalmanophyllum, 5-Densigrewingkia, 6-Grewingkia, 7-Crasilasma, 8-Streptelasma, 9- Dinophyllum, 10-Porfirieviella. See the explanation in the text.
Figure 2: Scheme development of Streptelasmatidae. The appearance of the corallites, nature of its sclerenchymal deposits, septum and interseptal camera: 1-Cothonion, 485,5 Ma, 2-Primitophyllum, 3-Kenophyllum, 4-Dalmanophyllum, 5-Densigrewingkia, 6-Grewingkia, 7-Crasilasma, 8-Streptelasma, 9- Dinophyllum, 10-Porfirieviella. See the explanation in the text.INFORMATION AND DISCUSSION ABOUT THE ORIGIN OF RUGOSA
Representatives Cothonion enough their features and characteristics, to consider them as credible ancestors rugosa and in particular the first of them, streptelasmatid. But more about that in the next paragraph.
Not everyone believes that rugosa, in the early stages of the evolution of cnidarians, connected its origin with coral genus Cothonion [3,17,18]. Nevertheless it facts and specific instances of these fossils rugosa remove existing doubts. Obviously, this was a small trochoid corals with characteristic rugosa deep cup, developed septa, fibrotic and trabecular microstructure corallites [8,10]. Corals possess bilateral symmetry and unique to rugosa to insert septal walls, which are clearly visible on the surviving corallites (Figure 3). They may be single thin plate shaped tabulae.
 Figure 3: Cothomionsimpomatum, a-c - Australia, b,d - Greenland. a,b - A septum are clearly visible and lateral line their insertion (to the left of the white line), c- clearly discernible structure wall and septum (indicated by arrows), typical for most Ordovitian and early Silurian Streptelasmatidae and Cystiphyllidae, d- lung bilateral symmetry corallite [8,10].
Figure 3: Cothomionsimpomatum, a-c - Australia, b,d - Greenland. a,b - A septum are clearly visible and lateral line their insertion (to the left of the white line), c- clearly discernible structure wall and septum (indicated by arrows), typical for most Ordovitian and early Silurian Streptelasmatidae and Cystiphyllidae, d- lung bilateral symmetry corallite [8,10].By the early Middle Ordovician in different seas and there rugosa Primitophyllum, Kenophyllum. If the first had a solid sclerenchymal thickening skeletal elements at all stages of growth, the presented Kenophyllumduring ontogeny markedly rid of these deposits. Interseptal camera in the early and middle stages of growth can not be traced. With age, they developed a coral, but remain rare, short and narrow. Tabulae, complete and rare, were appearing only in the later phases of ephebic growth stage (Figure 4). By the end of the Middle and Late Ordovician both species ceased to exist. More similar to rugosa characteristics not known to early Ordovician none of fossil animals.
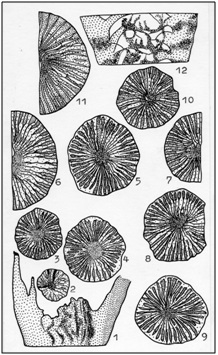 Figure 4: Ontogeny and variability Kenophyllumsubcylindricum Dybowski. 1-6, 7-9, 10-12 - multiple specimens of one layer, late Ordovician Siberian platform [1].
Figure 4: Ontogeny and variability Kenophyllumsubcylindricum Dybowski. 1-6, 7-9, 10-12 - multiple specimens of one layer, late Ordovician Siberian platform [1].Acceleration in ontogenesis sclerenchyma reduce some Kenophyllum by increasing the wok and the number of platelets led to the end of the first half Late Ordovician rugosa genera Dalmanophyllum These corals were more highly organized than their possible ancestors. Although they remained entirely with the early stages of merging septa without visible boundaries interseptal cameras and axis structure of sclerenchyma cover, but by the middle phase of neanic growth stage habitats most interseptal chambers partly exonerated from sclerenchyma deposits. At a later stage they were clearly distinguishable between each pair of baffles moderately incrassation of sclerenchyma.
Derived from Dalmanophyllum can be considered Densigrewingkia and Grewingkia as a result of further development of the ability of coral faster during growth to facilitate significant part of corallites abundant sclerenchyma.
Streptelasmatidae type Densigrewingkia just as their ancestors had, since the early stages may be exempted from part of sclerenchyma, but this release has been much more intense. Almost all interseptal cameras they differed with neanic growth stage. Further evolution of the tabulae, increase the convexity of the plates and their number, and the acceleration of the process of getting rid of sclerenchyme in ontogenesis leads to rugosa of genera Grewingkia. Majority Grewingkia from the earliest stages freed from a significant number of sclerenchyma, thereby facilitating corals. Many of these early differed interseptal camera. Densi Grewingkia exist until the end of the Ordovician, Grewingkia lived until middle Llandovery, Dalmanophyllum - until the end of Llandovery.
On the other hand, the development tendency streptelasmatid conversion axial structure, increasing the number and size of interseptal chambers thinning due to skeletal elements, large plates tabulae conversion into smaller convex, numerous, sometimes interseptal.
For corals with such an organization, first of all, are rugosagenera Crassilasma, which could occur from Kenophyllum, freeing up more skeletal elements and axial space coral formations on ample sclerenchyma. In the early stages of growth of these rugosa as well as that of their hypothetic ancestors were thick confluent septum without distinction interseptal cameras. The oldest Crassilasma keep a thickening neanic until the end, and even until middlebrephic growth stage. The majority of same Crassilasma ferns started to decline from the mid-to late neanic stage, revealing interseptal camera. But the final extinction, it is also like Kenophyllum, it did not occur even at the late stage of growth. All Crassilasma skeletal elements were thickened to a greater or sclerenchyma least coral throughout life, but in the axial space was not sclerenchyme solid. Septum, and filled if the axial zone, it is not in fuse structure. Reliable Crassilasma known since the beginning of the Late Ordovician, but their level of organization inner structure in the Late Ordovician says that the roots of these streptelasmatid should go into the Middle Ordovician, close to the time of occurrence and spread of Kenophyllum.
Strengthening the process of reducing sclerenchyme with septal apparatus and zoom interseptal cameras some Crassilasma could cause streptelasmatid genera Nelicelasma. These corals only in the very early stages of growth were abundant deposits of sclerenchyma. But from the middle phase neanic stage it began to decrease rapidly, facilitating the skeleton and labeling short interseptal chamber, the number and size of which increased rapidly with age coral. The axial zone filled with curled or variously curved, sometimes interrupted ends septum composing the porous structure of the centerline. On ephebic stage growth thickened skeletal elements were only peripheral parts of septum. This is significant progress in the development of single-band single corals to facilitate and improve the skeleton. Most of the life of a polyp was large enough and numerous interseptal camera and a relatively thin skeletal elements. Deputy Nelicelasma arose, probably in the second half Middle Ordovician and lasted until the end of the Late Llandovery.
Even more intensively going reduction in sclerenchyma corallites and above all, in the axial zone of the genus Streptelasma. These rugosa the earliest stage growth clearly differed interseptal cameras that with age, as well as skeletal elements, almost entirely absolve from sclerenchyma. With an average phase neanic stage growth most of these rugosa axial zone has remained free from any axial formations. At a later stage of growth continued to thicken sclerenchyma exiguous of the peripheral parts of the septum. Streptelasma existed since the middle of the Middle Ordovician to Late Llandovery.
With the passage of time to become a characteristic streptelasmatid only moderate thickening of the skeletal elements by sclerenchyma and development interseptal cameras free of it at an early growth habitats. The nature of the deposits sclerenchyma new feature appears in the process of evolution. During ontogeny, it is now not only reduced, but distributed differently in different parts of the coral. Septal apparatus of single-band rugosa obtain increasingly clear bilateral orientation, and the tabulae of the plate became more numerous, but smaller and more convex and counter sectors tend interseptal (Figure 5).
 Figure 5: Evolution of axial structures. 1-Kenophyllum, continuous filling of sclerechyme, 2-Helicelasma, grainy, perforation and thickening of the septum, 3-Crassilasma, axis scrolling fortek, continued growth of the septum, 4-Porfirieviella, and the absence of axial formations [1].
Figure 5: Evolution of axial structures. 1-Kenophyllum, continuous filling of sclerechyme, 2-Helicelasma, grainy, perforation and thickening of the septum, 3-Crassilasma, axis scrolling fortek, continued growth of the septum, 4-Porfirieviella, and the absence of axial formations [1].One of the first streptelasmatid with the development of the skeleton steel corals genera Dinophyllum. These were the early rugosa stage growth has considerably lightened the skeleton with moderate deposits sclerenchyma and clearly distinguishable inter septal cameras. With the growth of coral skeleton thickening reduces or distributed on the periphery of the septa around the perimeter of the cup, or only in the main and opposite sextants. Septal apparatus the end of the neanic stage, as a rule, acquired a clear bilateral orientation. Representatives Dinophyllum emerged, most probably from the Late Ordovician Crassilasma, reached its peak in Llandovery and, most likely, in early Wenlock ceased to exist.
Acquiring the ability to differentiate sclerenchyma with the aftermost early stages of growth could give some Kenophyllum to education streptelasmatid genera Pseudophaulactis.This coral extinction occurred first on skeletal elements opposite the top side, and then with middle growth stage began intensive thinning of the skeleton in the main sectors of the lower reclining side. In the later stages of growth sclerenchyma it disappeared completely or very slightly thickens peripheral ends of the septa, mostly cardinal directions (Figure 6) Pseudophaulactis known from the Late Ordovician to Late Llandovery.
 Figure 6: Ontogeny and variability Pseudophaulactislycophylloides Zapr. etIvnsk. 1-5,14, 6-13, 15-16 – multiple specimens of one layer, late Llandovery Siberian platform [1].
Figure 6: Ontogeny and variability Pseudophaulactislycophylloides Zapr. etIvnsk. 1-5,14, 6-13, 15-16 – multiple specimens of one layer, late Llandovery Siberian platform [1].Some Dinophyllum, Pseudophaulactis and Porfirieviella on late stages of growth in many ways reminiscent of primitive pleonophorn rugosa multiple thin septa, sometimes two cycles, raised numerous interseptal plates tabulae, clearly visible in the cross sections, minor deposits sclerenchyma vice peripheral rim. This is especially true for the latter species of corals.
OBSERVATION ON SYMMETRY RUGOSA
There is a concept of the origin Bilateria from Vendian bilaterally symmetrical coelenterates with metameric arrangement of the pockets of the gastric cavity. Ancestors Cnidaria attached to the substrate aboral pole, resulting in a reduction of the aboral nerve ganglion. Bilateral symmetry coral is treated as a characteristic feature of primitive cnidarians [20].
The fixed on the outer surface throughout the entire life cycle of the device orientation septal many rugosa demonstrates that it was bilaterally symmetrical animals (through the body and its coral skeleton can be carried out only one plane of symmetry). Metasepta insertion mechanism also shows that was inherent only rugosa bilateral symmetry [5,19,21-23]. Of all the possible ways of inserting the next successive partitions are "selected" only one - each next metaseptum inserted only in the direction of the main protosepta (Figure 7).
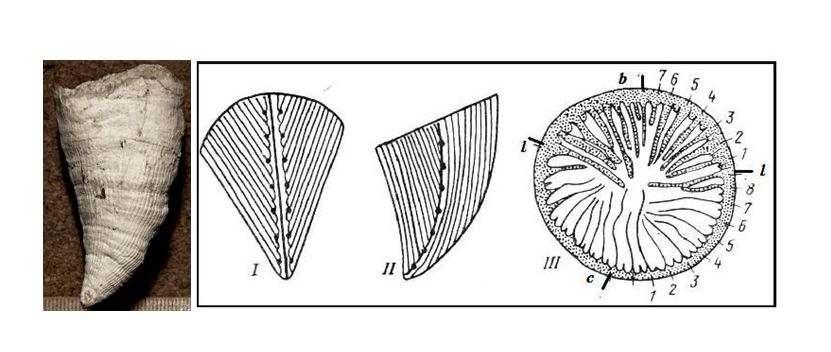 Figure 7: Inserting lines at the septum Streptelasmatidae: appearance (left) and (right).I- view from the main, II-side protosepta, III-in the cup: b, c, l - accordingly, basal (cardinal), opposite and side protoseptum, numerals marked metaseptum insertion sequence [1].
Figure 7: Inserting lines at the septum Streptelasmatidae: appearance (left) and (right).I- view from the main, II-side protosepta, III-in the cup: b, c, l - accordingly, basal (cardinal), opposite and side protoseptum, numerals marked metaseptum insertion sequence [1].Bilateral symmetry is hypothetical rugosa and was fixed skeleton themselves, so it should be formed much earlier, then there are, in the Precambrian. The principal types of invertebrates became isolated in Vendian and bilateral symmetry, such as trilobites is not in doubt. Qualitative restructuring of symmetry coelenterates on a completely different level of organization may have occurred in one of the critical boundaries of natural systems [8,25] at the beginning of the last milliardoletie.
If we assume that coelenterates - the first multicellular animals in Earth's history, and we know that 63-70% of species of multicellular Precambrian accounted for coelenterates, then we are entitled to assume that the first bilateral symmetry manifested in this group of animals.
CONCLUSION
REFERENCES
- Latypov YuYa (1984) The oldest solitary corals and their principles of Systematics. M.: Nauka, Page no: 94.
- Baars C, Pour MG, Atwood RC (2013) The earliest rugose coral. 150: 371-380.
- Scrutton CT (1999) Palaeozoic corals: their evolution and palaeoecology. Geology Today, 15: 184-193.
- Latypov YuYa (1977) Solitary cystifore corals North Asia. Novosibirsk, Nauka, Page no: 79.
- Latypov YuYa (1982) Solitary diafragma tophore corals of North Asia. M.: Nauka, Page no: 116.
- Schouppe A, Stacul P (1955) Die Genera Verbeekiella Penecke, Timorphyllum Gerth,Wannerophyllum n. gen., Lophophyltidium Grabauausdem Perm von Timor. - Palaeontographica A,.Suppl, 4, 5: 95-196.
- Iwanovwki AB (1975) RUGOSA. M.: Nauka, Page no:122.
- Jell PA, Jell JS (1976) Early Middle Cambrian corals from western New South Wales. Alcheringa An Australasian Journal of Palaeontology 1: 181-195.
- Scrutton CT (1997) The Palaeozoic corals, I: origins and relationships. Proceedings of the Yorkshire Geological Society 51: 177-208.
- Peel JS (2011) The coral Cothonion from the lower Cambrian of North Greenland. Alcheringa An Australasian Journal of Palaeontology 35: 405-411.
- Dzik J (1993) Early metazoan evolution and the meaning of its fossil record. Evolutionary Biology 27: 339-386.
- Linnaeus C von (17710 Mantissa Plantarum. Generumeditionis VI, etSpecierumeditionis II. Laurent Salvii, Stockholm 143-558.
- Fedorowski J (1997) Rugosa and Scleractinia - A commentary on some methods of phylogenetic reconstructions. Acta palaeontologtca polonica 42: 446-456.
- Hatschek B (1888)Lehrbuch der Zoologie. 1. Lieferung. Jena, Verlag von Gustav Fischer, 1-444.
- Haeckel E (1866) Generelle Morphologie der Organismen. Allgemeinegrundzüge der organischenformen-wissenschaft, mechanisch begründetdurch die von Charles Darwin reformirte descendenztheorie. Berlin, Verlag von Georg Reimer, Bd.1: 1-574; Bd.2: 1-462.
- Latypov YuYa (1975) Rugosa or Tetracoralla? Palaeontological Journal 3: 133-135.
- Engelbretsen M J (1993) A Middle Cambrian possible cnidarian coral from the Murrawong Creek Formation, NE New South Wales. Association of Australasian Palaeontologists Memoir, 15: 51-56.
- Fuller M, Jenkins R (2007) Reef corals from the Lower Cambrian of the Flinders Ranges, South Australia. Palaeontology, 50: 961-980.
- Milne-edwards H, Haime J (1850). A Monograph of the British Fossil Corals. London :Printed for the Palaeontographical society, Page no: 1850-1854.
- Malakhov VV (2004) The origin of bilaterally-symmetrical animals (BILATERIA). Zh Obshch Biol 65: 371-388.
- Hill D (1981) Treatise on Invertebrate Paleontology, Part F: Coelenterata. Supplement 1, Rugosa and Tabulata, volumes 1, 2. Boulder, Colorado: Geological Society of America; Lawrence, Kansas: University of Kansas Press,USA, Page no: 762.
- Wang G, Ian GP, Rongyu Li (2015) Remarks on the pattern of septal insertion in rugose corals. Alcheringa An Australasian Journal of Palaeontology 39: 1-39.
- Villee CA (1954) BIOLOGY. Saunders, Page no: 730
- Beklemishev VN (1964) Osnovy sravni tel’noianatomii bespozvonochnyh. Promolfologia. M.: Nauka,Page no: 433.
- Zhirmunsky AV, Kuzmin VI, Nalivkin VD, Sokolov BS (1980) Simulation of critical milestones in the development of systems and the periodization of history of the Earth. Vladivostok, 68. (in Russian) Sokolov, BS Fedonkin MA Early stages of the development of life on Earth. Cite ru-mo.
Citation: Latypov Y (2019) Some Reflections on the Beginning of the Oldest Corals Rugosa. Archiv Zool Stud 2: 008.
Copyright: © 2019 Yuri Latypov, et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.