The Role of Nerve Growth Factor in Atopic Dermatitis and the Modulation Effect of Pimecrolimus
*Corresponding Author(s):
Isaac MelamedImmunoe International Research Centers, Colorado, United States
Tel:+303 2244678,
Fax:+303 2244699
Email:melamedi@immunoe.com
Abstract
Atopic Dermatitis (AD) is a chronic, relapsing, inflammatory skin disorder. Recent studies indicate that AD is caused by a failure in signaling of apoptosis resulting from the dysregulation of T-cells and cutaneous tissue, with pivotal roles being played by Granzyme B and Nerve Growth Factor (NGF). Using cell lines and mononuclear cells from 20 atopic and non-atopic subjects, we examined the potential immune pathways that may lead to persistent inflammation as well as the role of pimecrolimus, an immunosuppressive/immune modulator agent, in treating AD. The results of the study indicate that low-dose pimecrolimus (1nM) up regulates the release of Granzyme B in CD4 and CD8 cells, thus stimulating apoptosis and healing. This enhanced apoptosis by pimecrolimus may clarify the mechanism of action and immune failure that leads to AD.
INTRODUCTION
Atopic Dermatitis (AD) is a chronic, relapsing, inflammatory skin disorder [1]. The inflammatory process seen in AD exhibits a mixture of immunological and pharmacological abnormalities that can be detected in serum and at the cellular level. Included in the immune histology of the skin of a person with atopic dermatitis are infiltrates of activated T-cells, dendrite presenting cells, eosinophils and mast cells.
Commencement of the immune cascade can result from multiple activators/activations, including allergens (food or environmental), infectious pathogens and a host of other distinctive triggers eg., stress or anxiety. This cascade often culminates in different biologic responses, including cell death. While programmed cell death, or apoptosis, can be triggered under a variety of circumstances, the mechanism by which cells are directed to cell cycle progression as opposed to apoptosis is not well understood. In recent years increasing evidence has suggested that regulatory T-cells play a pivotal role in the etiology of AD. The role of T-cells in the development of AD is complex and may involve various mechanisms [2].
Apoptosis plays an important role in skin inflammation. The cytokines perforin and granzyme B appear to have a strong correlation with the degree of skin inflammation and are important components in cell-mediated cytotoxicity [3]. Perforin, a plasma membrane, pore-forming protein, is part of lymphocyte cytotoxicity, which is mediated by lytic granules [4]. On activation, perforin and granzyme B are released through exocytosis and cooperatively trigger the death of their target cells (Figure 1).
Dysregulation of these compounds was recently demonstrated in patients with AD [3,5]. As T-regulatory cells are thought to be critically involved in skin inflammation, we hypothesize that cytotoxic granules released by skin infiltrating T-lymphocytes may be central to the disease process.
Pimecrolimus is an immunosuppressive/immune-modulator agent. The macrolide binds to macrophilin-12; the resulting complex then binds to the cytosolic and calcium/ calmodulin-dependent phosphatase calcineurin. This inhibits the ability of calcineurin to dephosphorylate the inactive form of nuclear factor of activated T-cells, ultimately inhibiting its translocation to the nucleus and its induction of type 1 (Tumor Necrosis Factor-alpha [TNF-α] Interferon-gamma [IFN-γ), type 2 (IL-4 and IL-5) and regulatory (Interleukin-10 [IL-10]) cytokines. Pimecrolimus also regulates activation of T-cells and mast cells, which may contribute to its anti-inflammatory effect [6,7]. In this study, we investigated the role of pimecrolimus in regulating some of the key components that play a part in atopic dermatitis.
METHODS
Part 1
A CD8-positive T-lymphocyte cell line, TALL, was used as a prototype “T-cytolytic” cell line. TALL requires passage in IL-2-supplemented culture media. Activation was performed as in the CD4 TRiPS line. A cloned human CD56-positive lymphocyte cell line, Natural Killer (NK-92), was used as a prototype “Natural Killer” cell line. NK-92 also requires passage in IL-2-supplemented growth media. Although these 3 cell lines have been adapted to in vitro culture conditions, they were not transformed (ie., not tumor) cells.
Three lymphocyte cell lines were separately pre-incubated in vitro ± NGF at a concentration of 125 pg/mL for 1 hour at 37?C, then incubated in titrated amounts of pimecrolimus, from 1, 10, 100, and 1000 nM, for an additional hour at 37?C. Cells (at a density of 2 x 105 in 1.0 ml Iscoves Modified Dulbeccos Minimal Essential Medium [IMDM] plus 10% Fetal Bovine Serum [FBS]) were then activated using suboptimal concentrations of either anti-CD3 plus anti-CD28 beads, a T-cell receptor complex (CD4- and CD8-positive lymphocyte lines) or cytokine stimulating factors (NK cell line) and incubated in vitro for an additional 18 hours at 37?C. Cell culture supernatants were then assayed for cytokine content and the cells were assayed for intracytoplasmic cytokines (granzyme B, perforin, IL-8, IL-16, INF-γ and TNF-α) by specific Enzyme Linked Absorbent Assay (ELISA). The presence of apoptosis was detected by staining cells from in vitro culture using “apo-direct” flow cytometric method to detect Deoxyribonucleic Acid (DNA) strand breakage in apoptotic cells.
A BioTek ELISA plate reader was used. ELISA kits for the following cytokines were utilized according to manufacturer protocols (suppliers are noted): Granzyme B (Bender Medsystems); Perforin (Cell Sciences); NGF (R & D Systems); IFN-γ, TNF-α, IL-8, IL-16 (Endogen).
Flow cytometry was performed on a Becton-Dickson FACSort flow cytometer using standard protocols. Antibodies utilized were: perforin (purchased from eBioscience) and Granzyme B, NGF, IFN-γ TNF-α , IL-8 and IL-16 (purchased from Caltag).
Part 2: Peripheral Blood Mononuclear Cells (PBMC) from human donors
Inclusion criteria for atopic subjects were mild to moderate AD involving 10% to 25% of body surface, with a severity score of 4 for any 3 of the 4 signs and symptoms (erythema, papulation, oozing/crusting, or lichenification). Subjects were excluded if they had used topical treatment for AD containing corticosteroids, antibiotics, or retinoids within 2 weeks and if systemic corticosteroids were used within 8 weeks. The use of systemic retinoids within 1 month was also exclusionary. Blood (10 cc) was drawn from each of the subjects; no further study procedures were performed. Plasma from donors was measured for NGF, granzyme B, perforin and TNF-α - using ELISA.
PBMC at 2 X 105 per 1.0 ml volume were isolated from the subjects with AD and from the non-atopic control donors; prepared by centrifugation on Ficol-Hypaque; and incubated ± pimecrolimus and ± NGF at 37?C for 1 hour. Cells were stimulated with 4 µg/ml Phytohemagglutinin (PHA) for 24 hours. Supernatants from cells were quantitated by ELISA for NGF, granzyme B, perforin, and TNF-α, while cells were assayed for apoptosis using APO-DIRECT methodology. Protocols for ELISA and flow cytometry were the same as those used in part 1 of the study.
STATISTICAL METHODS
Treatment effects were analyzed using repeated measures, including ANOVA, which takes into account correlation between subjects. Individual contrasts were calculated to compare treatment means of interest. All p-values were two-sided; all analyses were performed using SAS (SAS Institute, Inc., version 9.1).
RESULTS
Part 1: Human cell lines
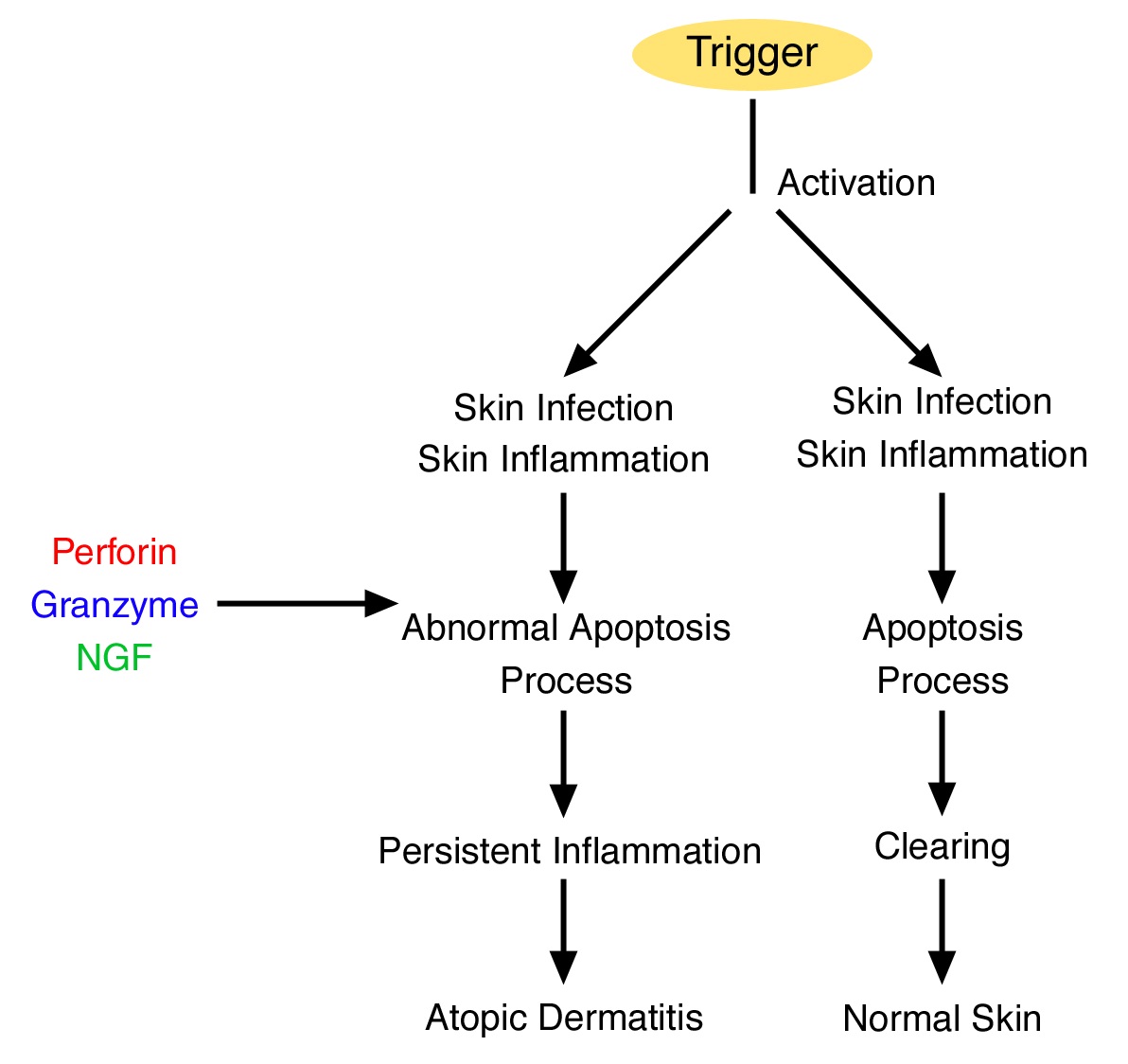
Figure 1: Role of perforin, granzyme B, and nerve growth factor in the atopic dermatitis cascade.
Stimulated CD4 cells incubated with NGF released 1514±198 pg/ml of granzyme B; the addition of 1nM pimecrolimus upregulated granzyme B release to 2878±461 pg/ml (p=0.0092). Pre-incubation of CD4 cells with NGF reduced upregulation of 1nM pimecrolimus on granzyme B from 3584±53 to 2878±461 pg/ml (p=0.058). Perforin release was the same in stimulated CD4 cells as in the presence of 1nM pimecrolimus (576±73, 514±37, respectively [p=0.26]). Perforin release in CD4 cells after incubation with NGF went from 576±73 to 367±112 pg/ml (p=0.54).
Pre-incubation of CD8-positive cells with 1nM of pimecrolimus increased granzyme B release from 1994±109 to 2706±21 pg/ml (p=0.0004). Pre-incubation with NGF slightly decreased the upregulation of granzyme B from 2153±47 to 2609±89 pg/ml (p=0.0014).
Pimecrolimus inhibited the release of IFN-γ, TNF-α and IL8 in CD4 cells. NGF augmented the inhibitory effect. IFN-γ and TNF-α remained unchanged in CD8 cells with 1nM pimecrolimus and NGF. IL-8 was inhibited in both CD4 and CD8 cells with pimecrolimus, and NGF augmented the inhibition.
IFN-γ gamma is a cytokine whose expression is stimulated from de novo messenger Ribonucleic Acid (mRNA) transcription. We were able to stimulate its production using anti-CD3+CD28. Pimecrolimus had a significant inhibitory effect at all tested concentrations (1 to 1000nM).
Pre-incubation of CD4 with 1nM pimecrolimus reduced the release of IFN-γ from 7253±173 to 5214±48 pg/ml (p<0.0001), while 10nM pimecrolimus reduced IFN-γ release to 1051±5.9 pg/ml (p< 0.0001).
Pre-incubation of CD4 cells with NGF augmented the inhibitory effect of 1nM pimecrolimus. IFN-γ release, with the addition of NGF, went from 7450±502 to 4712 ± 48 pg/ml after the addition of 1nM pimecrolimus (p=0.0007) (Figure 2).
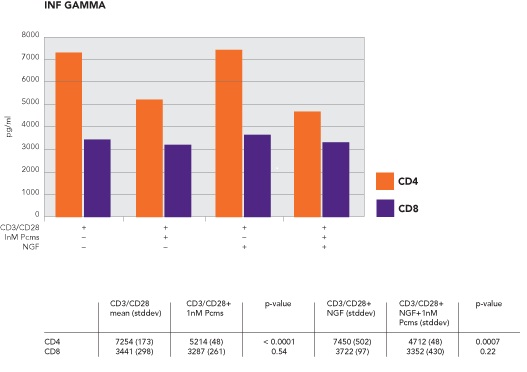
Figure 2: Effect of pimecrolimus on the release of IFN-? gamma.
In CD8 cells, 1nM pimecrolimus release of IFN-γ decreased from 3441±298 to 3287±261 pg/ml (p=0.54). After pre-incubation with NGF, IFN-γ levels decreased from 3722±97 to 3352±430 pg/ml (p=0.22) (Figure 3).
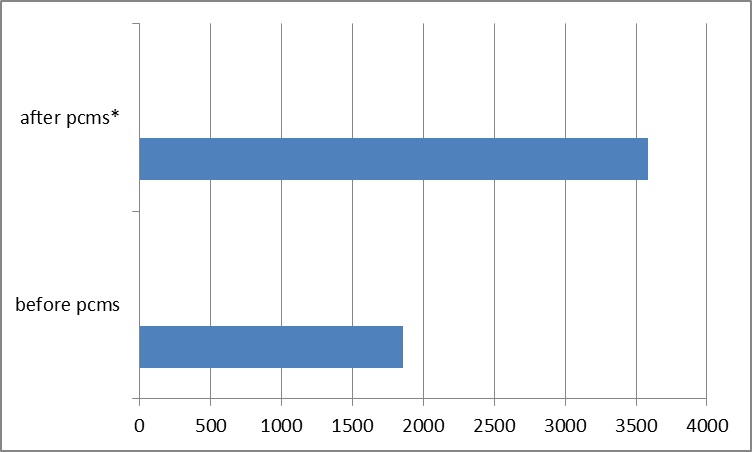
Figure 3: Effect of pimecrolimus on the release of granzyme B (pg/mL) *p<0.0001.
TNF-α is a cytokine whose expression is stimulated from de novo mRNA transcription. Its production was also induced by anti-CD3+CD28. Pimecrolimus at all concentrations tested (1-1000nM) produced significant inhibitory effect. In CD4 cells pre-incubated with 1nM pimecrolimus, TNF-α release was reduced from 420±4.7 to 154±7 pg/ml (p < 0.0001). At 10nM pimecrolimus, TNF-α release was reduced to 12±0.3 pg/ml (p<0.0001). Pre-culture with NGF produced a slightly significant inhibitory effect from 446±5.4 to 150±7.4 (p<0.0001) and 6±0.3 pg/ml (p<0.0001).
TNF-α release in CD8 cells was slightly increased in 1nM pimecrolimus, from 555±22 to 570±46 (p=0.67); this was not statistically significant. Higher doses of pimecrolimus, however, reduced the levels dramatically, 35±4.1, 13±1.4 (p<0.0001) and 13±1.4 pg/ml (p<0.0001). The addition of NGF had no effect at 1nM pimecrolimus: 558±15 to 554±21 pg/ml (p=0.80). Higher doses of pimecrolimus incubated with pimecrolimus followed the same trend.
IL-8 levels in CD4 and CD8 cells were inhibited with all pimecrolimus doses; NGF augmented the inhibition. IL-16 was not affected by pimecrolimus or NGF incubation.
IL-16 is a pre-formed cytokine in CD4-positive lymphocytes. A moderate background release was increased two-fold by stimulation with anti-CD3+CD28. Pre-incubation of CD4 T-cells with 1nM pimecrolimus left IL-16 release unchanged. Even with higher doses of pimecrolimus (10 and 100nM), there was no effect on IL-16 release. The same phenomenon was seen with NGF pre-incubation.
Pimecrolimus had no effect on granzyme B, perforin, IL-16, TNF-α and IL-8 release in NK cells, whereas pimecrolimus upregulated IFN-γ release in NK cells. IFN-γ release was upregulated at 1nM pimecrolimus, while NGF inhibited the upregulation.
Pre-incubation of NK cells up-regulated IFN-γ release with the addition of 1nM pimecrolimus, from 4916±132 to 7150±615 pg/ml (34%, p=0.004). Pre-incubation with NGF, however, inhibited upregulation of IFN gamma in NK cells (from 5213±598 to 6717±707 pg/ml) (7%, p=0.048).
Pimecrolimus reduced apoptosis in CD4 cells. NGF decreased the intensity of apoptosis. Activation of CD4 cells led to an increase in apoptosis (34%), compared to the nil control. Pre-incubation with pimecrolimus at any concentration resulted in findings 5.6+0.2% less than that observed with activation.
Pre-culture in NGF resulted in less intense apoptosis after stimulation of CD4 cells (mean log fluorescence intensity of 35 with NGF compared to 81 without NGF). The percentage of cells undergoing apoptosis, however, did not significantly change with NGF incubation (32%). In the presence of NGF, culture in any concentration with pimecrolimus had no additional effect on apoptosis (Figure 4).
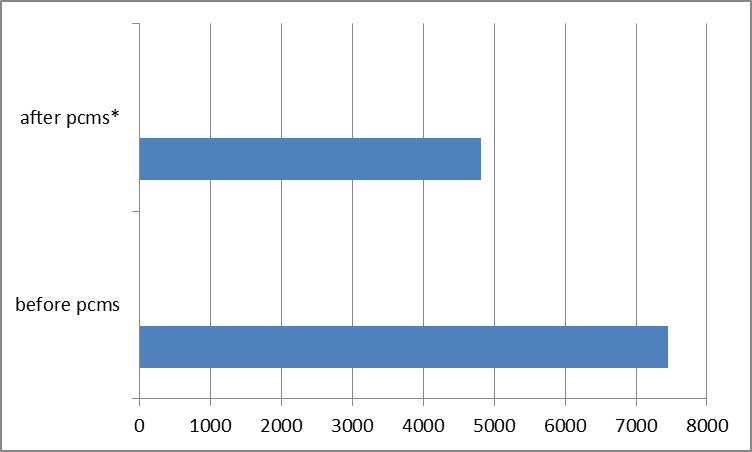
Figure 4: Effect of pimecrolimus on IFN-γ release in NGF augmented CD4 cells (pg/ml) *p=0.0007.
Part 2: PBMC human donor data
In non-atopic PBMC, the release of granzyme B was higher compared to atopic, 7452 compared to 6288 pg/ml (mean values). Pre-incubation with 1nM pimecrolimus had the same suppressive effect on granzyme B release in atopic subjects, compared to non-atopic, 51% and 50%, respectively, (p=0.0102).
Pre-incubation with NGF upregulated the release of granzyme B: in atopic subjects, from 6288 to 6672 pg/ml (7.6%), compared to non-atopic, from 7452 to 9146 pg/ml (23.6%, p=0.24). Levels of perforin and TNFα in the presence of pimecrolimus and NGF were extremely heterogeneous and non-predictable. Subject values were also calculated using natural logs, which gave similar results.
Apoptosis data
In atopic subjects, Phytohemagglutinin (PHA)-stimulated cells had a mean apoptosis of 9.46±9.8. With the addition of 1nM pimecrolimus, values were 12.3±9.9; this was not a significant change (p=.0906). The addition of NGF did not have an effect on apoptosis.
In non-atopic PHA-stimulated samples, apoptosis was 4.84±9:4; with the addition of 1nM pimecrolimus, the percentage increased to 10.2±13.0 (p=0.0542). The addition of NGF reduced the upregulation with PHA to 3.05±4.8. The addition of 1nM pimecrolimus further reduced the upregulation to 8.24±10.5 (p=0.0619).
Pimecrolimus (1nM) up regulates granzyme B release in CD4 and CD8 cells. Perforin is not affected, and NGF inhibits granzyme B release in both CD4 and CD8 cells.
Pre-incubation of CD4 positive T-helper cells with 1nM of pimecrolimus increased granzyme B release from 1860±33 to 3584±53 pg/ml (p<0.0001). Changes in perforin levels were not statistically significant. Pre-incubation of CD4 cells with 10nM of pimecrolimus had an inhibitory effect and reduced granzyme B release to 1572±51 pg/ml (p=0.0013). Reduction in perforin release was not statistically significant.
Stimulated CD4 cells incubated with NGF released 1514±198 pg/ml of granzyme B; the addition of 1nM pimecrolimus upregulated granzyme B release to 2878±461 pg/ml (p=0.0092). Pre-incubation of CD4 cells with NGF reduced the upregulation effect of 1nM pimecrolimus on granzyme B from 3584±53 to 2878±461 pg/ml (p=0.058). Perforin release was the same in stimulated CD4 cells as in the presence of 1nM pimecrolimus (576±73, 514±37, respectively [p=0.26]). Perforin release in CD4 cells after incubation with NGF went from 576±73 to 367±112 pg/ml (p=0.54).
Pre-incubation of CD8-positive cells with 1nM of pimecrolimus increased granzyme B release from 1994±109 to 2706±21 pg/ml (p=0.0004). Pre-incubation with NGF slightly decreased upregulation of granzyme B from 2153±47 to 2609±89 pg/ml (p=0.0014).
Pimecrolimus inhibited the release of IFN-γ, TNF-α, and IL8 in CD4 cells. NGF augmented the inhibitory effect. IFN-γ and TNF-α remained unchanged in CD8 cells with 1nM pimecrolimus and NGF. IL-8 was inhibited in both CD4 and CD8 cells with pimecrolimus, and NGF augmented the inhibition.
IFN-γ gamma is a cytokine whose expression is stimulated from de novo messenger Ribonucleic Acid (mRNA) transcription. We stimulated its production using anti-CD3+CD28. Pimecrolimus, at all concentrations tested (1 to 1000nM), had a significant inhibitory effect.
Pre-incubation of CD4 with 1nM pimecrolimus reduced release of IFN-γ from 7253±173 to 5214±48 pg/ml (p<0.0001), while 10nM pimecrolimus had a substantial inhibitory effect and reduced IFN-γ release to 1051±5.9 pg/ml (p< 0.0001).
Pre-incubation of CD4 cells with NGF augmented the inhibitory effect of 1nM pimecrolimus. IFN-γ release, with the addition of NGF, went from 7450±502 to 4712±48 pg/ml after the addition of 1nM pimecrolimus (p=0.0007) (Figure 2).
In CD8 cells, 1nM pimecrolimus release of IFN-γ decreased from 3441±298 to 3287±261 pg/ml (p=0.54). After pre-incubation with NGF, IFN-γ levels decreased from 3722±97 to 3352±430 pg/ml (p=0.22).
TNF-α is a cytokine whose expression is stimulated from de novo mRNA transcription. Its production was also induced by anti-CD3+CD28. Pimecrolimus at all concentrations tested (1-1000nM) produced significant inhibitory effect. In CD4 cells pre-incubated with 1nM pimecrolimus, TNF-α release was reduced from 420±4.7 to 154±7 pg/ml (p < 0.0001). At 10nM pimecrolimus, TNF-α release was reduced to 12 ± 0.3 pg/ml (p<0.0001). Pre-culture with NGF produced a slightly significant inhibitory effect at 1 and 10nM pimecrolimus, from 446±5.4 to 150±7.4 (p<0.0001) and 6±0.3 pg/ml (p<0.0001).
TNF-α release in CD8 cells was slightly increased in 1nM pimecrolimus, from 555±22 to 570±46 (p=0.67); this was not statistically significant. Higher doses of pimecrolimus, however, reduced the levels dramatically, from 35±4.1 to 13±1.4 (p<0.0001). The addition of NGF had no effect at 1nM pimecrolimus: 558±15 to 554±21 pg/ml (p=0.80).
IL-8 levels in CD4 and CD8 cells were inhibited with all pimecrolimus doses; NGF augmented the inhibition. IL-16 was not affected by pimecrolimus or by incubation with NGF.
IL-16 is a pre-formed cytokine in CD4-positive lymphocytes. A moderate background release was increased two-fold by stimulation with anti-CD3+CD28. Pre-incubation of CD4 T-cells with 1nM pimecrolimus left IL-16 release unchanged, from 158±8.1 to 140±16 pg/ml (p=0.16). Even with higher doses of pimecrolimus at 10 and 100nM, no effect was noted on IL-16 release (146±4.1 [p=0.08], 142±8.1 pg/ml [p =0.07] for 10nM and 100nM, respectively). The same phenomenon was seen with NGF pre-incubation (158±2.4 and 136±10 pg/ml for 10nM and 100nM pimecrolimus, respectively).
Pimecrolimus had no effect on granzyme B, perforin, IL-16, TNF-α and IL-8 release in NK cells, whereas pimecrolimus upregulated IFN-γ release in NK cells. IFN-γ release was upregulated at 1nM pimecrolimus. NGF inhibited the upregulation.
Pre-incubation of NK cells up-regulated IFN-γ release with the addition of 1nM pimecrolimus, from 4916±132 to 7150±615 pg/ml (34%, p=0.004). Pre-incubation with NGF, however, inhibited the upregulation of IFN gamma in NK cells (from 5213±598 to 6717±707 pg/ml) (7%, p=0.048).
Pimecrolimus reduced apoptosis in CD4 cells. NGF decreased the intensity of apoptosis process. Activation of CD4 cells led to an increase in apoptosis (34%), compared to the nil control. Pre-incubation with pimecrolimus at any concentration resulted in findings 5.6+0.2% less than that observed with activation.
Pre-culture in NGF resulted in less intense apoptosis after stimulation of CD4 cells (mean log fluorescence intensity of 35 with NGF compared to 81 without NGF). The percentage of cells undergoing apoptosis, however, did not significantly change with NGF incubation (32%). In the presence of NGF, culture in any concentration with pimecrolimus had no additional effect on apoptosis.
DISCUSSION
In this study, we explored the effect of pimecrolimus on perforin and granzyme B release and their effect on apoptosis. In analyzing the data from CD4 and CD8 cells, we noted that: 1) pimecrolimus at low doses (1nM) upregulated release of granzyme B; and 2) addition of NGF lowered upregulation. Furthermore, we discovered that the perforin levels were not changed, which did not fit with our original hypothesis.
IFN-γ and TNF-α levels were affected differently in CD4 and CD8 cells. These cytokines were not affected in CD8 cells but were inhibited in CD4 cells, with NGF augmenting the inhibition. IL-8 secretion was inhibited in both CD4 and CD8 cells, with NGF increasing the inhibition. IL-16 was not affected in either cell line. NK cells were unaffected by pimecrolimus and NGF in all cytokines, except for IFN-γ at low doses (1nM) where it was upregulated; NGF reduced upregulation.
In subject data, our analysis focused on PBMC, which contains all monocyte cells. This may explain the heterogeneous and non-predictable values for perforin and TNF-α. The release of granzyme B in the sample population was reduced in atopic subjects compared to non-atopic subjects. Our hypothesis that atopic dermatitis could be linked to a dysregulation of granzyme B release correlates with this result. Our perforin results, however, could not be correlated to our hypothesis, possibly due to the small sample size, the mixture of cells in PBMC, or due to the different signaling pathway associated with perforin or granzyme B.
NGF inhibited granzyme B release, showing that it is involved in the mechanism of AD. Apoptosis trends showed an increase with the addition of 1nM pimecrolimus in both atopic and non-atopic subjects.
In atopic dermatitis, T-cells and an altered immune defense play roles in the development of compromised epithelial function and clinical disease. Atopy may be the prerequisite for failure of the immune cascade and may prepare the “right” culture for immune-related diseases in later stages. The regulation of both normal and allergic immunity is fully T-cell dependent and relies exclusively on the activation and action of different subtypes of T-cells and their products, which in turn activate and direct the entire immune system network and involved tissue cells.
Patients with AD have increased susceptibility to certain bacterial, viral and fungal infections. There is strong evidence to support the role of Staphylococcus aureus infection in the pathogenesis of AD [1]. Additionally, staphylococcal super-antigen can facilitate keratinocyte presentation of allergens to CD4 T-cells, thus establishing a link between 2 of the most common AD diseases. Perforin and granzyme B are strongly expressed in lesional skin and show significant correlation with apoptotic keratinocytes. Disorders of regulatory T-cells also appear to predispose patients to dermatitis and an allergic phenotype, as shown by the IPEX syndrome, which is associated with mutation within FoxP3, a critical transcription factor for regulatory T-cells [8]. Affected individuals may have autoimmune enteropathy, dermatitis, food allergy, and/or other autoimmune phenomena. In addition to the infectious role, allergens themselves may indirectly mediate T-cell influence on barrier function and may also have a direct effect. For example, Der=p1 has been shown to directly influence tight structure junction by its protease [2].
A major pathway by which T-cells clear infection is the targeted exocytosis of granules containing various cytolytic proteins, including the pore forming proteins, perforin (pfp) and members of the granzyme serine protease (gsp) family towards the infected cells [9]. Once delivered, pfp and gsp act synergistically to initiate programmed cell death by both caspase-dependent and independent pathways.
The most well characterized cytolytic effector protein is granzyme B, which cleaves substrate proteins, resulting in classic apoptotic death characterized by mitochondrial depolarization, nuclear and DNAase activation. Given its important role in mediated killing, many infectious agents have evolved strategies that target and disable granzyme B function. The atopic patients in our study displayed “disabled systems” when granzyme B was involved.
In this study, we showed that, upon T-cell activation, granzyme B release in atopic subjects is reduced when compared to non-atopic subjects. Moreover, we upregulated the release of granzyme B with lower doses of pimecrolimus. Upregulation of granzyme B evidenced with pimecrolimus was not seen with regards to perforin and was not observed in all T cell populations studied. We did not see this upregulation in NK cells, suggesting that: 1) pimecrolimus has a selective effect on various T-cell groups; and 2) the pathways that regulate granzyme B pathways may differ from the perforin pathways.
Whether the trigger is infectious or allergenic, the immune cascade may lead to cell activation or cell apoptosis. Persistent inflammatory processes have been evaluated as a failure of an apoptosis process. Efficient immune system function can clear apoptotic cells, while persistent inflammation is testimony of a failure within the immune system itself. This was shown in this study when T cells, either in cell lines or isolated from atopic subjects, were not able to efficiently release granzyme B and, thus, clinically presented with AD due to accumulation of apoptotic cells.
Based on the cell line and donor data, we have, for the first time, shown that low-dose pimecrolimus upregulates granzyme B release. Upregulation of granzyme B can be part of a major mechanism that enhances the apoptosis process and allows for healing.
Various cells are involved in the allergic ‘symphony.’ Eosinophils appear to be key players [10]. A recent study suggests that pimecrolimus inhibits eosinophilic superoxide production and Eosinophil Derived Neurotoxic (EDN) [11]. This suggests that, although various pathways are regulated by pimecrolimus, there is a common final endpoint: regulation of apoptosis signaling. Pimecrolimus does not have the same upregulation effect on perforin as on granzyme B.
The functional role of NGF and its receptors in allergic diseases is still an object of investigation [12]. Previous studies have demonstrated that NGF has an active role in allergic inflammation [13]. Based on our previous work, this role is shared between the nervous and immune regulatory cells [14]. In this study, we confirm our hypothesis regarding the role of NGF in allergic skin inflammation. Moreover, we suggest that NGF, which may play an important role in the mechanism of AD: 1) can regulate the release of granzyme B; 2) plays a key role in the apoptosis cascade; and 3) as NGF, may amplify keratinocyte-mast cell interaction and enhance skin inflammation through the interplay between structural and immune system. There is growing evidence that NGF is a signaling molecule supporting inflammation [15,16]. NGF may play a central role in the pathogenesis of AD and pimecrolimus may interfere in the interplay of NGF and immune cells.
CONCLUSION
The rationale for this study was based on recent data proposing that atopic dermatitis is an immunological process resulting from the inability of T cells or cutaneous tissue to be normally regulated due to a failure in the signaling of apoptosis. As a consequence of this failure, patients lose the immunological barrier that may have later immunological consequences such as asthma development.
The results of this study indicate that low-dose pimecrolimus (1nM) upregulates granzyme B release in CD4 and CD8 cells, which can enhance the apoptosis process and have a positive effect on the healing process.
ACKNOWLEDGEMENT
The authors thank Irene Durham, MS, for editorial support on this manuscript.
FUNDING SOURCES
Novartis Pharmaceuticals Corporation provided funding support.
AUTHOR CONTRIBUTIONS
Conceived and designed the experiments: Isaac Melamed, David Bar-Or, Richard Shimonkevitz
Analysed the data: Isaac Melamed, David Bar-Or, Richard Shimonkevitz, Lacey Robinson, Melinda Heffron
Wrote the first draft of the manuscript: Isaac Melamed, Melinda Heffron, Lacey Robinson
Contributed to the writing of the manuscript: Isaac Melamed, David Bar-Or, Richard Shimonkevitz, Lacey Robinson, Melinda Heffron
Agree with manuscript results and conclusions: Isaac Melamed, David Bar-Or, Richard Shimonkevitz, Lacey Robinson, Melinda Heffron
Jointly developed the structure and arguments for the paper: Isaac Melamed, David Bar-Or, Richard Shimonkevitz, Lacey Robinson, Melinda Heffron
Made critical revisions and approved final version: Isaac Melamed, David Bar-Or, Richard Shimonkevitz, Lacey Robinson, Melinda Heffron
All authors reviewed and approved the final manuscript.
REFERENCES
- Leung DYM (2000) Atopic dermatitis. Allergy Immunol 105: 860.
- Ogg G (2009) Role of T cells in the pathogenesis of atopic dermatitis. Clin Exper Allergy 39: 310-316.
- Yawalkar N, Schmid S, Braathen LR, Pichler WJ (2000) Perforin and granzyme B may contribute to skin inflammation in atopic dermatitis and psoriasis. Br J Dermatol 144: 1133-1139.
- Stepp SE, Matthew PA, Bennett M, de Saint Basile G, Kumar V (2000). Perforin: more than just an effector molecule. Immunol Today 21: 254-256.
- Melamed I, Franklin RA, Brodie C, et al. (1993) Signaling of B-lymphocytes by nerve growth factor: tyrosine phosphorylation of phospholipase Cg1, MAP2-Kinase and gp140trk. Molec Biol Cell 4: 123.
- Grassberger M, Baumruker T, Enz A, Hiestand P, Hultsch T et al. (1999) A novel anti-inflammatory drug, SDZ ASM 981, for the treatment of skin diseases: in vitro pharmacology. Br J Dermatol 141: 264-273.
- Arndt KA, et al. (2001) Tacrolimus and pimecrolimus. Sem Cutaneous Med Surg. 4: 215-281.
- Chantila TA (2008) Molecular Mechanisms if Regulatory T-cell Development. In: Blazer K (ed.). T-cell Regulation in Allergy, Asthma and Atopic Skin Diseases. Clinical Immunology and Allergy, Karger Publishing, Switzerland. Vol 94.
- Trapani JA, Smyth MJ (2002) Functional significance of the perforin/granzyme cell death pathway. Nat Rev Immunol 2: 735-747
- Dombrowicz D, Capron M (2001) Eosinophils, allergy and parasites. Curr Opin Immunol 13: 716-720.
- Plager DA, Henke SA, Matsuwaki Y, Madaan A, Squillace DL et al. (2009) Pimecrolimus reduces eosinophil activation associated with calcium mobilization. Int Arch Allergy Immunol 149:119-126.
- Bresciani M, Lalibertè F, Lalibertè MF, Gramiccioni C, Bonini S (2009) Nerve growth factor localization in the nasal mucosa of patients with persistent allergic rhinitis. Allergy 64: 112-117.
- Rochlitzer S, Nassenstein C, Braun A (2006) The contribution of neurotrophins to the pathogenesis of allergic asthma. Biochem Soc Transactions 34: 594-599.
- Melamed I (2009) The role of Zytalin in neuro-inflammatory diseases: Nerve-growth factor as a possible link between the nervous and immune system [abstract]. Presented at the 45th Annual Meeting of the Drug Information Association (DIA), June 23, 2009 San Diego, CA, USA.
- Nockher WA, Renz H (2006) Neurotrophins and asthma: Novel insight into neuroimmune interaction. J Allergy Clin Immunol 117: 67-71.
- Nockher WA, Renz H (2006) Neurotrophins in allergic diseases: From neuronal growth factors to intercellular signaling molecules. J Allergy Clin Immunol 117: 583-589.
Citation: Melamed I, Robinson L, Heffron M, Bar-Or D, Shimonkevitz R (2016) The Role of Nerve Growth Factor in Atopic Dermatitis and the Modulation Effect of Pimecrolimus. J Clin Dermatol Ther 3: 021.
Copyright: © 2016 Isaac Melamed, et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.