Biotechnology of Accelerated Breeding and Improvement of Cotton Varieties
*Corresponding Author(s):
Shadmanov RKInstitute Of Genetics And Plants Experimental Biology Of The Academy Of Sciences Of Uzbekistan, Uzbekistan
Tel:+7 (99893) 1849654,
Email:Rshad@yandex.ru
Abstract
The success of any breeding program depends upon the availability of genetic variability of traits of interest and availability of efficient markers for selection of traits. Within the last 20 years, molecular biology has revolutionized conventional breeding techniques in all areas. Biochemical and molecular techniques have shortened the duration of breeding programmes. In this article are discussed the opportunities of use of biochemical markers (enzymes and proteins) for acceleration of breeding of new and improving of existing cotton varieties on tolerance to different unfavourable environmental factors and with complex of desired traits and varietal purity. We analyzed nineteen (19) local cotton varieties and lines (Gossypium hirsutum L.) for this purpose. The indices of oxidoreductases class enzymes and two phosphoprotein markers were developed as objective biochemical markers of earliness, homogeneity, tolerance to Verticillium wilt, drought and salinity.
Keywords
Biochemical markers; Cotton; Isoenzymes; MAS; Micro evolutionary processes; Plant breeding; Proteins; Tolerance; Unfavourable environmental factors
INTRODUCTION
In most cases, plants of agricultural crops influenced by various unfavorable factors show stability to these factors, as a result of the adaptation. It happens mainly by means of active adaptations and micro evolutionary processes in populations of plants [1,2].
It should be emphasized that the leading role in the maintenance of intracellular homeostasis and adaptation to stressors is played by enzymes [3]. Thereby the induction of enzymes or elements of their qualitative structure with new properties, or new proteins providing cells protection is an essential factor of survival. It is pointed out that isoenzyme systems possess undesirable for markers ontogenetic, tissue or organoid specificity, variability depending on environmental factors [4]. While absolute advantage of the isozymes is that they are not neutral genome markers [5]. Taking part in different biochemical processes and physiological reactions, they determine organism’s adaptation to various environmental factors [6,7]. They have monogenic genetic control, are co-dominant in comparison with some types of DNA markers. In addition, their resolving power is high enough [8].
Using indices of seed enzymes and proteins we have bred cultivars and lines of cotton with such characteristics as earliness, yielding capacity, tolerance to diseases, tolerance to drought and salinity for extremely short period of time.
MATERIALS AND METHODS
Plant material
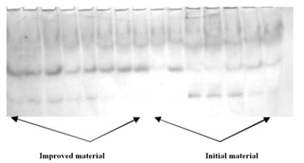
Figure 1: Selection for tolerance to Verticillium wilt on phenoloxidase AN-Bayaut-2 cotton variety.
SELECTION OF ADAPTIVE GENOTYPES
Phosphoproteins extraction
Germination of selected embryos and generation of seedlings
RESULTS AND DISCUSSION
Resistance to salinity
We have suggested that the question of tolerance to salinity could be connected with protein structures and their components’ metabolism process. Providing one or another configuration in cell elements, these protein structures protect these cell elements against excessive invasion of salt.
While differentiated analysis of seeds proteins we have detected two phosphor proteins in seeds of resistant to salinity cotton plants (Figure 2). In seeds of non-resistant cotton plants these proteins were not detected. Being objective marker traits, these proteins allow selecting genotypes from plants population (Figure 3) and determine level of tolerance to salinity of any cotton cultivar. It’s been found out, that two phosphoproteins, typical for tolerant to salinity genotypes, were proteins with relative electrophoretic mobility (Rf)=0.37 and (Rf)=0.56
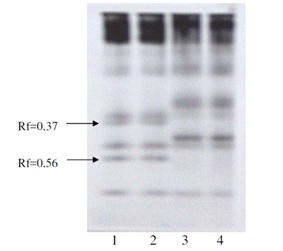 Figure 2: Marker for tolerance to salinity. (Proteins of phosphoproteins fraction).
Figure 2: Marker for tolerance to salinity. (Proteins of phosphoproteins fraction).Note: 1 and 2 - tolerant to salinity biotypes, 3 and 4 - non-tolerant to salinity biotypes
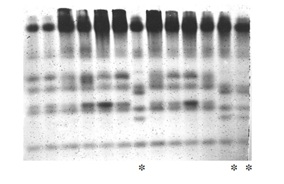 Figure 3: Selection for tolerance to salinity in population of cotton plants Namangan-77 cotton variety.
Figure 3: Selection for tolerance to salinity in population of cotton plants Namangan-77 cotton variety.Tolerant to salinity biotypes
 Figure 4: a) Genotype received after selection from Khorezm-127 variety. b) Initial variety Khorezm-127 (Control check).
Figure 4: a) Genotype received after selection from Khorezm-127 variety. b) Initial variety Khorezm-127 (Control check).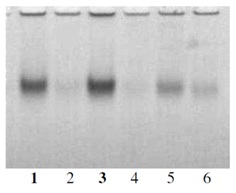 Figure 5: Marker for tolerance to drought and salinity.
Figure 5: Marker for tolerance to drought and salinity.1-Shodlik-11; 2-Shodlik-1; 3-Shodlik-9; 4-Tashkent-6; 5-108-F; 6-Bukhara-6.1, 3-tolerant to salinity cultivars.
|
Cotton Varieties |
% of Non Resistant to Drought Plants |
|
C-4534 |
70.4 |
|
Shodlik-11 |
18.8 |
|
C-6524 |
35 |
According to the field experiments, plants of the experimental, resistant to water deficit Shodlik-11 cotton variety, exceeded non-resistant to water deficit plants in growth and development after selection. And this difference was approximately 15-20%.
Resistance to Verticillium wilt
Uzbekistan is one of the northern most cotton growing areas in the world. The climate in Uzbekistan provides favorable conditions for Verticillium wilt (Verticillium dahliae) infection, and yield losses can exceed 40% [14,15]. A role of a polyphenols-polyphenoloxidase system in immunity of plants was well known. Attempts to treat the plants with gossypol to decrease fungal damage have also been unsuccessful. The reason was that high activity of polyphenoloxidase has not been connected to immunity or mitochondria’s respiratory system that produces ??P.
Penetration of pathogen in a plant occurs already on the initial stage of germination. Pathogen should pass through epidermal layer cells of the seed coat or young root [14]. In case of presence of active enzymatic systems, oxidizing processes occur intensive in this layer. Products of oxidation possess stronger fungicide activity, in comparison with initial substances and form a powerful chemical barrier [16].
Many researchers observed, but could not explain, mass accumulation of fungi on the surfaces of a root and it’s insignificant penetration into plants tissue of plants with low vulnerability to disease and almost complete absence of fungi on roots, but with high content of infection in tissues of susceptible plants. Meeting on the way of penetration cells with high enzymatic activity, the infection eliminates (immune genotypes) or penetrates in vascular system of plant weakened or in insignificant amount. It does not greatly influence the plant’s normal developmental processes (tolerant genotypes). We came to conclusion that two non-mitochondrial respiratory pathways, non producing ??P are responsible for resistance to wilt infection. The terminal oxydases of these pathways are phenol- and ascorbatoxidases. Analyzing tissues of plants we have found out that not all genotypes reveal presence of non mitochondrial systems of metabolism. In resistant to wilt genotypes they were always at amount, which provides immunity. It has been shown that phenoloxidase works during initial period of germination of seeds, and ascorbatoxidase-during the period of budding and flowering. The progeny of the genotypes selected according to the signs of these enzymes had a high level of tolerance to pathogens in comparison with initial plants. Thus, from the experiments, phenoloxidase enzyme can serve as an effective biochemical marker for both the tolerance to wilt infection (Verticillium dahliae) (Figure 1 and Figure 6) and for varietal purity evaluation (Figure 7).
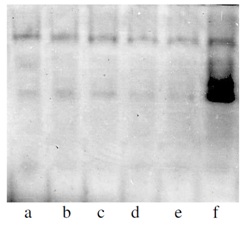 Figure 6: Marker for tolerance to wilt (Verticillium dahliae) (Polyphenoloxidase)
Figure 6: Marker for tolerance to wilt (Verticillium dahliae) (Polyphenoloxidase)A-E - Non-tolerant to Verticillium wilt genotypes
F- Tolerant to Verticillium wilt genotype
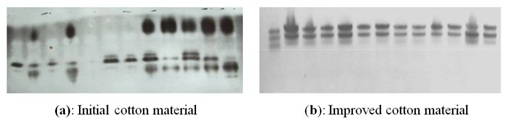 Figure 7: Evaluation of varietal purity (Phenoloxidase).
Figure 7: Evaluation of varietal purity (Phenoloxidase).We have analyzed the genotypes from the population of susceptible to Verticillium wilt AN Bayaut cultivar for tolerance to wilt (Verticillium dahliae) during the research (Figure 7). The analysis of the results showed, that the cotton material improved by means of marker-assisted selection with use of biochemical characteristics of enzymes exceed the initial cotton material on tolerance to Verticillium wilt and was homogeneous. According to the field experiments, tolerant to Verticillium wilt plants exceeded non-tolerant. The results meet the table data (Table 2). Both L-2 and Shodlik-9 variety were received from the original susceptible to wilt AN-Bayaut-2 variety, and Line-3 was received from the standard C-4727 variety. According to the table, damage from wilt of the experimental L-2, L-3, Shodlik-9 lines and varieties received, were significantly lower in comparison with initial material.
|
Cotton lines and cultivars |
% of Infected plants |
|
Namangan-77 (Standard check) |
20 |
|
AN-Bayaut-2 (Original susceptible cultivar) |
62 |
|
Shodlik-9 |
3.5 |
|
L-2 |
3.6 |
|
?-4727 (Original susceptible cultivar) |
85 |
|
L-3 |
4.6 |
In order to successfully develop cotton and other crops cultivars, traditional breeding methods and phenotypic screening need to be combined with new approaches and technologies including biochemical and DNA markers assisted Selection to make a breeding process more accurate and efficient. It should be emphasized, that incontrovertible advantage of DNA and protein markers is, that they are closer, then other substances to or are the carrier of hereditary information themselves (DNA) [17].
Plants unlike animals due to the fact, that they are tightly bound to the habitats, during long term evolution and changing environmental conditions, had developed a wide range of defense mechanisms. In the process of plants domestication the necessity in some of these defense mechanisms eliminated, but they was preserved in genome [18]. And there are always biotypes with such mechanisms in plants population. But phenotypic selection, especially towards unfavorable environmental factors, is not always efficient to develop these genotypes.
Under adaptation we usually understand a long process in which many generations are involved; however, within a population individual plants can make useful adaptive changes within one generation. Biochemical adaptation takes place on different levels of the metabolism by replacement of amino acids in primary sequence of enzyme, or by change of isoenzymes balance, which cause the proteins polymorphism [19].
The micro evolutionary processes are one of the major factors causing genetic polymorphism of natural populations including cultivars. They lead to occurrence of genetically various individuals in population that finds reflection in distinctions of members of a population by inherited diversified traits: morphological, physiological, biochemical, qualitative and quantitative [20]. But the problem is being able to identify the few useful individuals among the entire population of plants. Often the trait is not visually detectable or varies depending on environmental conditions or presence of a pest. Therefore, selection by the phenotype of plants, especially in relation to pathogens, is not always efficient or is time consuming because of the random factor. Beside for that, it is necessary to take into account that microorganisms also adapt. But, biotypes which have arisen as a result of adaptation, maintain high tolerance to pathogens much longer. The biotechnology, based on identification of such genotypes on marked traits allows very quickly, already at a level of seeds conducting selection for many traits particularly or in complex.
Selection for earliness
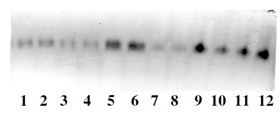 Figure 8: Selection for earliness (catalase).
Figure 8: Selection for earliness (catalase).5, 6, 9, 12-Genotypes selected for earliness.
Together with increasing of tolerance to Verticillium wilt and reduction of vegetative period we have increased weight of one boll and oil content in seeds (Table 3 and 4).
|
Plant Characteristics (Standard) |
Namangan-77 (Standard) |
??-16 |
??-18 |
Shodlik-3(9) |
III-5 |
Shodlik-10 |
|
|
Plant height, (centimeter) |
108.5±0.4 |
110.0±0.2 |
115.0±0.2 |
105.0±0.2 |
100.0±0.1 |
115.0±0.2 |
|
|
Number of bolls |
14.0±0.1 |
20.0±0.2 |
20.0±0.2 |
19.0±0.1 |
17.0±0.09 |
20.0±0.1 |
|
|
Resistance to wilt disease, % of healthy |
45.0±0.3 |
98.5±0.2 |
96.0±0.2 |
99.0±0.1 |
98.0±0.2 |
98.5±0.1 |
|
|
Yield capacity, Hwt/hectare |
30 |
38 |
37 |
40 |
37.5 |
36.5 |
|
|
Fiber length, mm |
33.0±0.2 |
34.5±0.19 |
37.5±0.2 |
34.5±0.2 |
34.5±0.1 |
35.5±0.3 |
|
|
Fiber output, % |
37.0±0.17 |
37.5±0.2 |
37.6±0.2 |
38.5±0.23 |
37.5±0.19 |
37.0±0.1 |
|
|
Micronair, mic |
4.8±0.14 |
4.2±0.1 |
4.1±0.2 |
4.1±0.2 |
4.4±0.1 |
4.3±0.1 |
|
|
Oil content in seeds, % |
19.5±0.2 |
22.5±0.3 |
24.0±0.3 |
25.5±0.2 |
23.5±0.2 |
23.0±0.2 |
|
Cotton Lines and Varieties |
Days to Maturity |
No of Bolls on a Plant |
One Boll Weight |
Lint % |
1000 Seed |
|
Namangan-77 (standard check) |
108 |
13.2±0.15 |
5.3±0.1 |
38.2±0.1 |
115±0.3 |
|
Bayaut-2 (original susceptible variety) |
114 |
12.1±0.1 |
5.5±0.09 |
35.7±0.1 |
115±0.3 |
|
Shodlik 9 |
108 |
15.2±0.1 |
7.4±018 |
38.7±0.1 |
141±0.2 |
|
Line-2 |
108 |
14.0±0.12 |
7.0±0.1 |
38.5±0.12 |
139±0.2 |
|
C-4727(Original susceptible variety) |
113 |
13.1±0.2 |
6.0±0.17 |
36.2±0.1 |
125±0.17 |
|
Line-3 |
110 |
12.5±0.2 |
6.6±0.2 |
38.6±0.2 |
125±0.2 |
CONCLUSION
Biochemical markers correlating with such important traits as tolerance to drought and salinity, tolerance to wilt, earliness, evaluation of variety’ purity and morphological homogeneity were developed and tested with experiments in this research. On their basis we received and tested in field genotypes with complex of above traits. As for resistance to Verticillium wilt, the most simple and effective was electrophoretic quantitative and qualitative assessment of oxidase respiration systems. In particular, regarding tolerance to wilt it was shown that use of isozymes indices of phenoloxidase on seed material was effective for selection of adaptive genotypes.
As for tolerance to salinity we found out that the tolerance was connected to the synthesis of structural proteins and their components, which through their configuration in cells, prevent penetration of the salt. In the course of the study on reserve seed proteins was found out that seed of cotton resistant to high salinity levels always contained two phosphor-proteins that were not present in non- resistant genotypes. Isozymes indices of the glucose-6-phosphate dehydrogenase enzyme were also effective for selection for salinity resistance. According to the results of the experiments with a number of dehydrogenase enzymes we found out, that glucose-6-phosphate dehydrogenase enzyme can serve as indirect marker for simultaneous estimation and selection for two traits, tolerance to drought and salinity.
The research showed that seeds’ isozymes analysis allows not only developing genetic predisposition on different traits, but also traits correlation. For example, the study of earliness trait of cotton plants showed that the most essential trait, which could be combined with earliness was tolerance to wilt (Verticillium dahliae), because early ripening cultivars are mainly non-tolerant to wilt infection, or faster lose their tolerance during reproduction. According to the study results, catalase enzyme was identified as an indicator for development of early ripening genotypes. And this is not due to coincidence as catalase forms a part of the metabolic respiratory system that defines and controls the level of activity for various metabolic processes.
Thus, the research results showed that it is possible to develop physiologically and Biochemically adapted biotypes in cultivars populations. Using biochemical markers on seed allozymes level, we were able to develop genotypes with tolerance to unfavorable factors and to increase homogeneity and cultivar Population’ purity. Such markers based on biological polymers’ polymorphism, could be unique phenotypic markers reflecting allelic state of corresponding genes. Their discretization appearing stable on seed level and being environment independent allows high accuracy identification of genotypes, and also allows accelerating breeding process.
ACKNOWLEDGEMENT
REFERENCES
- Deveroll BJ (1980) Plants defence mechanisms. Pg no: 126 [In Russian].
- Harward G (1985) Introduction to ecological biochemistry. Mir Publishers, Moscow, Russia, [In Russian].
- Hochachka PW, Somero GN (1988) Biochemical adaptation. Mir Publishers, Moscow, Russia. Pg no: 568 [In Russian].
- Rick CM (1982) lsozymes in plant breeding. California Agriculture 36: 28.
- Tanksley SD, Orton TJ (1983) Isozymes in plant genetics and breeding, (1st edn). Elsevier, Amsterdam, Netherlands.
- Sejdimirova OA, Yanbaev YuA, Zaytsev DYu (2009) Isozyme markers in research of variability of spring wheat varieties, recognized in Bashkortostan. OSU Bulletin No. 6.
- Wijsman HJW, Petunia (1983) Isozymes in plant genetics 406 and breeding, Part B. Elsevier, Amsterdam, Netherlands.
- Ivachenko LE, Lavrent’yeva SI, Konichev AS, Golokhvast KS (2016) The role of enzymes in the adaptation of soybean of different philogenetic origin to growing conditions. Der Pharma Chemica 8: 236-244.
- Shields CR, Orton TJ, Stuber CW (1983) An outline of general resource needs and procedures for the electrophoretic separation of active enzymes from plant tissue. Developments in Plant Genetics and Breeding 1: 443-468.
- Ivachenko LE (2008) Methods of studying soybean enzyme polymorphism. Blagoveshchensk State Pedagogical University, Blagoveshchensk, Russia.
- Wendel JF, Weeden NF (1989) Visualization and interpretation of plant isozymes. In: Soltis DE, Soltis PS (eds.). Isozymes in plant biology. Dioscorides Press, Portland, USA.
- Shadmanov RK, Igamberdiyeva DI (1996) Patent No. UZ 3100 Method of cotton plants’ salt tolerance determination.
- Hasegava PM, Bressan RA, Zhu JK, Bohnert HJ (2000) Plant cellular and molecular responses to high salinity. Annu Rev Plant Physiol Plant Mol Biol 51: 463-499.
- Bell AA (1992) Verticillium wilt. In: Hillocks RJ (ed.). Cotton Diseases. C.A.B. International, Wallingford, UK.
- Bell AA (1995) Mechanisms of disease resistance in Gossypium species and variation in Verticillium dahliae. In: Constable GA, Forrester NL (eds.). Challenging the Future: Proceedings of World Cotton Research Conference 1. CSIRO, Melbourne, Australia.
- Avazkhodjaev MH, Zeltser SSh (1980) Physiological factors of cotton tolerance to wilt. Tashkent Pg no: 45-64.
- Konarev AV, Konarev VG, Gubareva NK (2000) Seed proteins as markers in solution to a problem of genetic sources of plants. Breeding and seed growing// Cytology and genetics. 34: 91-104 [In Russian].
- Metlitskiy LV, Ozertskovskaya OL (1985) How plants defense themselves against diseases. Mir Publishers. Pg no: 190 [In Russian].
- Rubin BA, Artsikhovskaya EV (1963) Biochemistry and physiology of plant immunity. Pergamon Press, London, UK.
- Rider CC, Taylor CB (1980) Isoenzymes. Chapman and Hall, London, UK.
Citation: Shadmanov RK, Shadmanova AR, Scheffler JA (2018) Biotechnology of Accelerated Breeding and Improvement of Cotton Varieties. J Biotech Res Biochem 1: 001.
Copyright: © 2018 Shadmanov RK, et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.