Herbivory and Soil Properties as Ecological Constraints to Willow Growth in a Restored Riparian Ecosystem in Coastal California
*Corresponding Author(s):
Gretchen CoffmanDepartment Of Environmental Management, University Of San Francisco, San Francisco, California, United States
Tel:+1 3106001901,
Email:gccoffman@usfca.edu
Abstract
This study investigated stunted arroyo willow (Salix lasiolepis) cuttings planted along a restored riparian corridor in Marin County, California. Restoration activities along this corridor included channel realignment, bank stabilization, and placement of streambed material and large woody debris, which required the use of heavy machinery and compacted soils. Additionally, evidence of deer browsing indicated that predation maybe significantly affecting cutting growth and survivorship. In our field study, we measured effects of deer browsing and physical soil characteristics (i.e., soil moisture, compaction, texture, and total organic content) on the change in willow growth (i.e., height, mean canopy diameter, estimated aerial percent cover, and volume) of 160 recently planted willow cuttings (in their second and third year of growth following planting) to better understand stunted growth. Growth and health attributes of the cuttings were further analyzed in relation to three factors: 1) presence of exclusionary deer fencing; 2) location on left or right bank and 3) age of the cutting. Results of the study indicated deer herbivory was a critical stressor limiting survivorship and growth of willow cuttings. Exclusionary fencing resulted in significantly higher growth for all four metrics compared to unfenced willows with the greatest beneficial effect on younger willows in their second growing season. Significantly higher soil moisture and lower soil compaction were found in reference sites compared to restoration study sites. However, the influence of physical soil characteristics on willow growth revealed weak correlations. Exclusionary fencing is a cost-efficient method for restricting browsing by wild herbivores on recently planted willow cuttings, most effective when implemented during the first two growing seasons or until willow cuttings are resilient to the effects of herbivory.
Keywords
INTRODUCTION
Active revegetation of native riparian species is a critical component of most riparian restoration projects. Native woody riparian species typically grow quickly and support the ecosystem by stabilizing streambanks [1], increasing root density for erosion prevention [2], and establishing channel vegetation structure for faunal habitats [3]. Woody riparian species provide valuable ecosystem services by increasing retention of flood water, reducing sedimentation, regulating temperature by providing shade, and improving water quality [4,5]. Willows (Salix spp.) are commonly utilized for revegetation of riparian ecosystems due to their simple propagation, demonstrated high survivorship, and rapid growth rate. Willows can reach heights of over 10 meters (m) and may grow approximately 2 to 3 m after a single growing season, thereby rendering most branches inaccessible for browsing by herbivores [6]. While their rapid growth makes willows resistant to browsing, cuttings are highly susceptible to intense browsing during the first few years after being planted [6]. Excessive grazing has been shown to negatively affect plant growth and reproduction [7-9], completely eradicate populations of woody plants [10], and has contributed to the worldwide degradation of riparian ecosystems [11,12]. Over time, frequent removal of woody stems and foliage by herbivores can result in a reduction in the carbon reserves below ground surface, thereby preventing natural defense mechanisms, including chemical resistance and rapid vertical growth, rendering the plants more susceptible to browsing [13].
Disturbed soil conditions can also adversely affect willow growth and overall survivorship. Construction activities commonly required for riparian restoration projects include: channel realignment; bank stabilization; reconstruction of geomorphic features such as floodplain benches, riffle pools, and depositional basins; placement of streambed material and large woody debris; and grade control structures. The use of heavy machinery and intensive ground disturbance required for these restoration activities can result in heavily compacted soils and reduced quantities of topsoil and organic material. Although increased soil compaction reduces the potential for soil erosion, it can also reduce the total porosity of the soil [14], effectively decreasing the potential for water to infiltrate the soil. Soil compaction may also reduce carbon and nitrogen cycling, soil microbial biomass [15,16], or impede plant growth, specifically root length, depth, and penetration through the soil [17]. Stunted willow growth may be indicative of an ecological stress which could negatively influence the overall success of a restoration project if not resolved. Due to the dynamic complexity of ecosystems, any number of ecological factors (e.g., excessive grazing by livestock, natural predation, poor soils, or low water availability) could detrimentally affect vegetation health [18].
Based on existing site conditions observed in the restored riparian ecosystem, we hypothesized that the leading ecological constraint contributing to stunted willow growth was either browsing by large herbivores or decreased soil moisture due to increased soil compaction resulting from ground-disturbing restoration activities. The objectives of our study were to 1) evaluate the effects of browsing by large herbivores on willow cuttings through the use of exclusionary fencing and 2) evaluate the effects of soil characteristics on willow growth through the collection of soil compaction, moisture, and composition data and comparison with data from a healthy riparian reference ecosystem.
METHODS
Study location
Experimental design
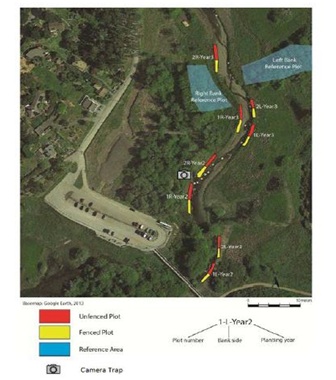 Figure 1: Location of restoration and reference plots along Redwood Creek at Muir Beach, California.
Figure 1: Location of restoration and reference plots along Redwood Creek at Muir Beach, California.|
Willow Age |
Bank |
Plot Number |
Total Number of Unfenced Replicates per Plot |
Total Number of Fenced Replicates per Plot |
|
Second Season |
Right |
1 |
10 |
10 |
|
Second Season |
Right |
2 |
10 |
10 |
|
Second Season |
Left |
1 |
10 |
10 |
|
Second Season |
Left |
2 |
10 |
10 |
|
Third Season |
Right |
1 |
10 |
10 |
|
Third Season |
Right |
2 |
10 |
10 |
|
Third Season |
Left |
1 |
10 |
10 |
|
Third Season |
Left |
2 |
10 |
10 |
|
|
Total number of cuttings |
80 |
80 |
|
Willow growth and herbivory
Soil characteristics
Although many natural processes contribute to compacted soils [19], natural causes typically only affect the top 5 cm of soil, whereas anthropogenic influences may affect soil 20 to 60 cm below ground surface (bgs) [17]. We sampled soil compaction using a soil compaction meter at approximate depths of 15, 30, 45, and 60 cm bgs, recording measurements in pounds per square inch (psi). Due to the variability between soil horizons, study site conditions, and the accuracy of the instrument, we categorized soil compaction readings as: 1) 0-200 psi; 2) 200-250 psi; 3) 250-300 psi; 4) 300-450 psi; and, 5) greater than 450 psi. A larger range was assigned to soil compaction ‘Category 1’ due to the relative ease at which the compaction meter penetrated soft soils and the inability to record static values in soil horizons that lacked significant resistance. We were unable to quantify soil compaction greater than 450 psi due to extreme compaction and instrument limitations.
We also measured soil texture and organic matter within the study plots. For each study plot, we collected and analyzed eight composite soil samples composed of seven subsamples consisting of cores collected at a depth of 45 cm bgs. Following thorough mixing of the subsamples, a single composite sample was extracted for each of the eight study plots. Soil samples were dried and total organic content (TOC) was calculated following the Environmental Protection Agency’s protocol for loss-of-ignition [20]. We determined grain size and soil texture using a hydrometer.
Reference plots
Statistical analysis
Since we collected willow growth data at the beginning and end of the growing season, we averaged the monthly soil moisture and soil compaction data at each of the 80 cuttings to illustrate the representative soil characteristics over the course of the growing season. We converted the categorical data to continuous data by using the average of the categorical ranges. We conducted two-way ANOVA for soil moisture and soil compaction to determine if seasonal averages of each study plot were related (Systat Software, Version 13). We also compared the monthly average soil moisture and soil compaction of the study plots to the monthly averages of the reference plots. We then paired the monthly averages of soil moisture and compaction data with the growth differences in height and canopy cover for each cutting. Using pairwise Pearson r-correlation analysis, we analyzed each soil characteristic against each growth metric, investigating for any potential correlations, isolating cutting age and bankside for each pairwise analysis.
RESULTS
|
Factors of Interest |
|||||||
|
Growth Metrics |
Bank |
Year |
Fenced |
Bank*Year |
Bank*Fenced |
Year*Fenced |
Bank*Year*Fenced |
|
Height |
F [1,150] = 2.536 |
F [1,150] = 97.115 |
F [1,150] = 37.351 |
F [1,150] = 11.572 |
F [1,150] = 27.631 |
F [1,150] = 29.345 |
F [1,150] = 7.27 |
|
Canopy Diameter |
F [1,150] = 10.943 |
F [1,150] = 28.587 |
F [1,150] = 62.105 |
F [1,150] = 31.721 |
F [1,150] = 11.335 |
F [1,150] = 8.624 |
F [1,150] = 18.368 |
|
Percent Cover |
F [1,125] = 11.861 |
F [1,125] = 4.705 |
F [1,125] = 16.974 |
F [1,125] = 0.004 |
F [1,125] = 5.671 |
F [1,125] = 8.969 |
F [1,125] = 4.059 |
|
Volume |
F [1,123] = 13.655 |
F [1,123] = 54.811 |
F [1,123] = 26.469 |
F [1,123] = 3.055 |
F [1,123] = 18.674 |
F [1,123] = 24.370 |
F [1,123] = 6.093 |
Legend: * = 0.05 ≥ P > 0.01 = significant, ** = 0.01 ≥ P > 0.001 = highly significant, *** = P ≤ 0.001 = very highly significant based on a 95% confidence level.
Willow height
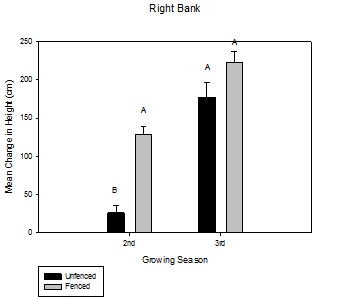 Figure 2: Mean change in height over the growing season between fenced and unfenced and second-year versus third-year willow cuttings on the right bank.
Figure 2: Mean change in height over the growing season between fenced and unfenced and second-year versus third-year willow cuttings on the right bank.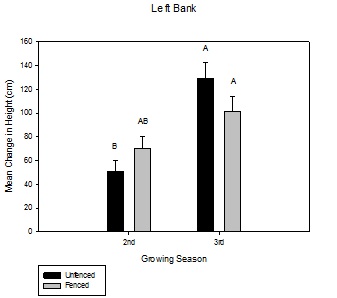 Figure 3: Mean change in height over the growing season between fenced and unfenced and second-year versus third-year willow cuttings on the left bank.
Figure 3: Mean change in height over the growing season between fenced and unfenced and second-year versus third-year willow cuttings on the left bank.Willow canopy diameter
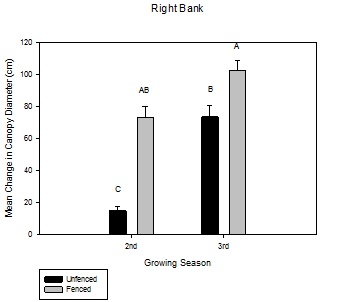 Figure 4: Mean change in canopy diameter (cm) over the growing season between fenced and unfenced and second-year versus third-year willow cuttings on the right bank.
Figure 4: Mean change in canopy diameter (cm) over the growing season between fenced and unfenced and second-year versus third-year willow cuttings on the right bank.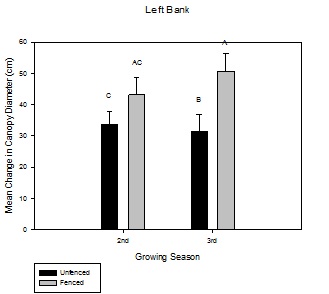 Figure 5: Mean change in canopy diameter (cm) over the growing season between fenced and unfenced and second-year versus third-year willow cuttings on the left bank.
Figure 5: Mean change in canopy diameter (cm) over the growing season between fenced and unfenced and second-year versus third-year willow cuttings on the left bank.Willow aerial percent cover
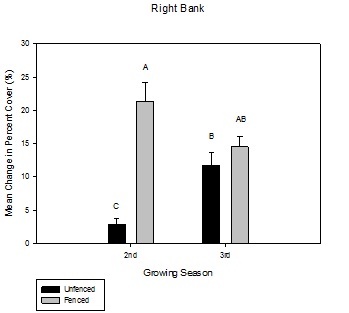 Figure 6: Mean change in percent aerial cover over the growing season between fenced and unfenced and second-year versus third-year willow cuttings on the right bank.
Figure 6: Mean change in percent aerial cover over the growing season between fenced and unfenced and second-year versus third-year willow cuttings on the right bank.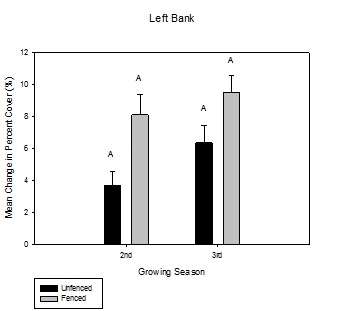 Figure 7: Mean change in percent aerial cover over the growing season between fenced and unfenced and second-year versus third-year willow cuttings on the left bank.
Figure 7: Mean change in percent aerial cover over the growing season between fenced and unfenced and second-year versus third-year willow cuttings on the left bank.Willow volume
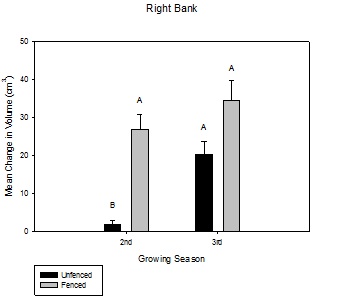 Figure 8: Mean change in volume (cm3) over the growing season between fenced and unfenced and second-year versus third-year willow cuttings on the right bank.
Figure 8: Mean change in volume (cm3) over the growing season between fenced and unfenced and second-year versus third-year willow cuttings on the right bank.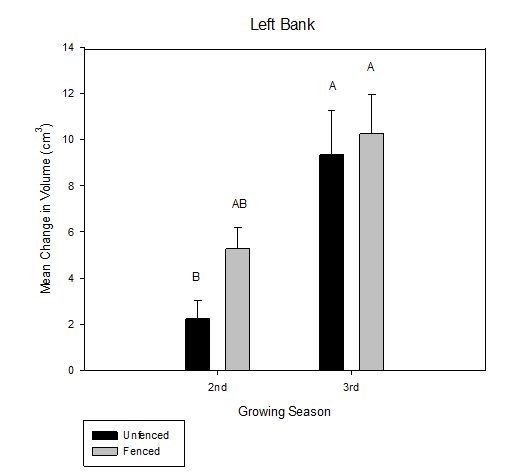 Figure 9: Mean change in volume (cm3) over the growing season between fenced and unfenced and second-year versus third-year willow cuttings on the left bank.
Figure 9: Mean change in volume (cm3) over the growing season between fenced and unfenced and second-year versus third-year willow cuttings on the left bank.Deer herbivory
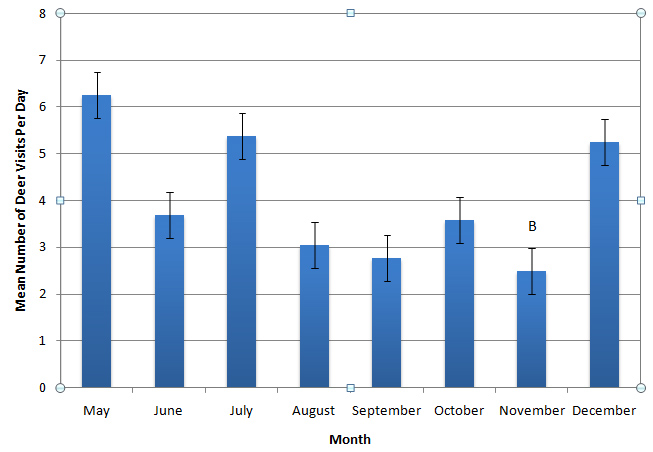 Figure 10: Mean number of deer visits per day for unfenced willows in their second growing season.
Figure 10: Mean number of deer visits per day for unfenced willows in their second growing season.Soil characteristics
Soil moisture varied significantly between months, plots, and cuttings in restoration and reference sites. Soil compaction levels remained relatively consistent throughout the study period. Soil probes were capable of penetrating the ground surface with minimal effort within the reference plots. However, soils within the study plots were so heavily compacted that the soil compaction meter was not capable of penetrating to the first sample depth of 15 cm. Soil moisture and compaction for the reference plots indicated very strong inverse correlations at all depths (r-values ranging from -0.764 to -0.963), with increased soil moisture in less compact soils. Similarly, we observed many strong correlations between soil moisture and compaction within the study plots (Table 3). Analysis of the six composite samples identified similar soil characteristics within the study and reference plots; however, the study plot samples contained approximately half of the quantity of TOC (4.1 to 5.0 percent by volume) compared to the reference plot samples (7.6 and 10.9 percent).
Soil characteristics vs willow growth
|
Study Plot Type |
Bank |
Grain Size |
Texture |
Organic Matter (% by volume) |
||
|
Sand (%) |
Silt (%) |
Clay (%) |
||||
|
Third Season |
Right |
31.75 |
47.00 |
21.25 |
Loam |
4.6 |
|
Third Season |
Left |
34.5 |
44.25 |
21.25 |
Loam |
4.1 |
|
Second Season |
Right |
45.00 |
41.25 |
18.75 |
Loam |
4.9 |
|
Second Season |
Left |
60.75 |
25.50 |
13.75 |
Sandy Loam |
5.0 |
|
Reference |
Right |
30.5 |
48.25 |
21.75 |
Loam |
10.9 |
|
Reference |
Left |
55.5 |
30.75 |
13.75 |
Sandy Loam |
7.6 |
DISCUSSION
Our analysis of soil characteristics concluded that restoration activities created significantly different soil conditions within the restored portions of the study area compared to undisturbed reference site conditions. Significant differences in soil moisture and compaction were observed between the study and reference plots, with mean soil moisture within the study plots measured at nearly half that of the soil within the reference plots and mean soil compaction measured at more than double that of the soil within the reference plots. These variations could be attributed to the difference in distance from the stream or the difference in soil texture between our study and reference plots [21].
Evette et al. [22] found that all growth metrics of willow cuttings are significantly related to available soil moisture and drought conditions generally increase willow growth stress compared to moist, well-drained conditions [23,24]. Highly-compacted soils may result in increased soil strength, reduced bulk density, and a reduction in water-holding capacity, all of which can restrict root growth and hinder plant health [25]. Furthermore, natural recovery of compacted soils may require many years depending on the regional climate, soil texture, and the level of compaction. Contrary to findings of similar studies, we only found weak correlations between soil characteristics and willow growth.
Our study identified a clear effect of willow growth within the exclusionary fencing along the banks of Redwood Creek. This effect was especially well pronounced in the second growing season. Fenced willow cuttings generally exhibited increased growth and increased biomass compared to cuttings located outside of fenced plots during the growing season. These results indicate that grazing contributed to stunted growth of planted willow cuttings. Although many studies have found a correlation between unrestricted browsing by herbivores and adverse effects to vegetation growth, the findings of our study were unique in that the effect of fencing was different between second- and third-year cuttings and the sides of the bank.
Overall, most fenced second-year cuttings demonstrated an increased change in growth for all metrics compared to unfenced cuttings. However, fenced third-year cuttings demonstrated no significant difference in growth metrics compared to unfenced third-year cuttings. These results suggest that the effects of browsing have a greater detrimental effect on second-year willows compared to third-year willows in the same habitat. Other studies have also found the effects of herbivory to depend on the age of the willows [26-29]. Several studies have shown that the nutritional value of willow leaves and shoots in their first two years of growth is valuable to large herbivores [29,30]. Additionally, juvenile willows are likely to be more accessible to herbivores due to their smaller stature and may have reduced chemical defenses as they prioritize early rapid growth [27]. Therefore, it may be possible that willows at Redwood Creek were not only more accessible to herbivores but may have also been higher in nutritional value if they continued to prioritize the production of photosynthetic material over chemical defenses.
The effectiveness of exclusionary fencing between second- and third-year willows suggests resilience to the effects of herbivory was different between the two age groups. These results differ from studies which conclude that younger willows may demonstrate increased compensatory responses to herbivory than older willows due to greater accumulation of stored reserves. However, Hanley and Fegan [31] found that younger willows were more susceptible to the detrimental effects of herbivory since biomass removal can greatly reduce the reserves necessary for compensatory responses. Therefore, younger willows with less biomass may have a reduced ability to recover under conditions of unrestricted browsing. Our results indicate that willow cuttings at this site become more resistant to the effects of herbivory by the third growing season following initial planting; however, the mechanism (chemical or reserves) for resistance remains unknown.
Willows are a common preferred food source for herbivores in riparian ecosystems [32]. Observations of increased growth among willows protected from browsing in this study were consistent with many studies, including De Jager et al. [33], who demonstrated improved survivorship and growth among willow communities following the removal of large herbivores. However, several studies used fencing to restrict access by livestock, not wild herbivores [34,35]. Our study corroborates findings from other studies that the use of exclusionary fencing can be a successful tool for protecting restored vegetation from wild herbivores as well as livestock [28,36].
Our study suggests that the use of exclusionary fencing may be an effective method for facilitating recovery of riparian vegetation at riparian restoration sites where herbivory is prevalent. Understanding the timing and frequency of browsing is particularly valuable information for installing exclusionary fencing treatment [37]. Fencing has a greater impact if implemented during the first two years following initial planting when willows are most accessible by herbivores, or until willows demonstrate sufficient tolerance to effects of herbivory. High levels of browsing found in our study may be more detrimental during the growing season because the tearing of leaves directly affects the ability of the plant to photosynthesize; whereas during the winter, browsed willows have demonstrated increased shoot growth, branch frequency, and bud formation [38].
While the use of exclusionary fencing to restrict access by large herbivores and reduce the stress of browsing proved to be an effective method for protecting newly planted willow cuttings in this study, it is not a universally applicable solution and may not be appropriate for all restoration projects. Depending on site conditions, fencing could result in additional adverse effects such as inadvertent entrapment of wildlife or obstruction of wildlife corridors. In this study, fenced plots were originally intended to remain in place for at least two consecutive years; however, during the winter season, water levels rose substantially in Redwood Creek. Due to concerns that fencing could inadvertently trap fish utilizing the riparian corridor, fencing was removed before final measurements were taken for the remainder of the wet season and re-installed in April. This is why we did not show results for first year willow cuttings. While the removal of exclusionary fencing was not anticipated, it was a necessary action and should be taken into consideration for similar riparian restoration projects. Therefore, while the use of exclusionary fencing can be an effective, low-cost, low-maintenance method for restricting browsing by large herbivores; it is important to consider the potential indirect effects of implementing this method.
ACKNOWLEDGEMENT
The authors gratefully acknowledge the NPS and GGNRA for allowing us to conduct this study at Redwood Creek throughout ongoing restoration efforts. We greatly appreciate the University of San Francisco, Environmental Science Department and Masters of Science in Environmental Management Program.
REFERENCES
- Gray DH, Barker D (2004) Root-soil mechanics and interactions, water science and application. American Geophysical Union, Washington, D.C., USA.
- Wynn TM, Mostaghimi S, Burger JA, Harpold AA, Henderson MB, et al. (2004) Ecosystem restoration: Variation in root density along stream banks. Journal of Environmental Quality 33: 2030-2039.
- Young JA, Clements CD (2003) Seed germination of willow species from a desert riparian ecosystem. Journal of Range Management 56: 496-500.
- Bernhardt ES, Sudduth EB, Palmer MA, Allan JD, Meyer JL, et al. (2007) Restoring rivers one reach at a time: Results from a survey of U.S. river restoration practitioners. Restoration Ecology 15: 482-493.
- Allan CJ, Dosskey MG, Vidon P, Gurwick NP, Allan CJ, et al. (2010) The role of riparian vegetation in protecting and improving chemical water quality in streams. Journal of the American Water Resources Association 46: 261-277.
- Bergström R, Guillet C (2002) Summer browsing by large herbivores in short-rotation willow plantations. Biomass Bioenergy 23: 27-32.
- Morris WF, Hufbauer RA, Agrawal AA, Bever JD, Borowicz VA, et al. (2007) Direct and interactive effects of enemies and mutualists on plant performance: A meta-analysis. Ecology 88: 1021-1029.
- Chun YJ, van Kleunen M, Dawson W (2010) The role of enemy release, tolerance and resistance in plant invasions: Linking damage to performance. Ecol Lett 13: 937-946.
- Oduor AMO, Gomez JM, Stauss SY (2010) Exotic vertebrate and invertebrate herbivores differ in their impacts on native and exotic plants: A meta analysis. Biological Invasions 12: 407-419.
- Olofsson J, Kitti H, Rautiainen P, Stark S, Oksanwn L (2001) Effects of summer grazing by reindeer on composition of vegetation, productivity and nitrogen cycling. Ecography 24: 13-24.
- Robertson AI, Rowling RW (2000) Effects of livestock on riparian zone vegetation in an Australian dryland river. Rivers Research and Applications 16: 527-541.
- Jansen A, Robertson IA (2001) Relationships between livestock management and the ecological condition of riparian habitats along an Australian floodplain river. Journal of Applied Ecology 38: 63-75.
- Barto EK, Rillig MC (2010) Does herbivory really suppress mycorrhiza? A meta-analysis. Journal of Ecology 98: 745-753.
- Silva AP, Barros N, Costa L, Leite F (2008) Soil compaction and eucalyptus growth in response to forwarder traffic intensity and load. Revista Brasileira de Ciência do Solo 32: 921-932.
- Frey B, Kremer J, Rudt A, Sciacca S, Matthies D, et al. (2009) Compaction of forest soils with heavy logging machinery affects soil bacterial community structure. European Journal of Soil Biology 45: 312-320.
- Pupin B, Silva F, Nahas E (2009) Microbial alterations of the soil influenced by induced compaction. Revista Brasileira de Ciéncia do Solo 33: 1207-1213.
- Nawaz MF, Bourrié G, Trolard F (2013) Soil compaction impact and modelling. A review. Agronomy for Sustainable Development 33: 291-309.
- Sunil C, Somashekar RK, Nagaraja BC (2011) Impact of anthropogenic disturbances on riparian forest ecology and ecosystem services in Southern India. International Journal of Biodiversity Science, Ecosystem Services & Management 7: 273-282.
- Drewry JJ (2006) Natural recovery of soil physical properties from treading damage of pastoral soils in New Zealand and Australia: A review. Agricultural Ecosystem Environment 114: 159-169.
- Schumacher BA (2002) Methods for the determination of Total Organic Carbon (TOC) in soils and sediments. United States Environmental Protection Agency (EPA), Environmental Sciences Division National Exposure Research Laboratory, Las Vegas, Nevada, USA.
- Dinesh K, Bansal ML, Phogat VK (2009) Compactability in relation to texture and organic matter content of alluvial soils. Indian J Agric Res 43: 180-186.
- Evette A, Labonne S, Rey F, Liebault F, Jancke O, et al. (2009) History of bioengineering techniques for erosion control in rivers in Western Europe. Environ Manage 43: 972-984.
- Lower SS, Orians EM (2003) Soil nutrients and water availability interact to influence will growth and chemistry but not leaf beetle performance. Entomologia Experimentalis et Applicata 107: 69-79.
- Greer E, Pezeshki SR, Shields FD (2006) Influence of cutting diameter and soil moisture on growth and survival of black willow salix nigra. Journal of Soil and Water Conservation 61: 311-323.
- Souch CA, Martin PJ, Stephens W, Spoor G (2004) Effects of soil compaction and mechanical damage at harvest on growth and biomass production of short rotation coppice willow. Plant Soil 263: 173-182.
- Guillet C, Bergstrom R (2006) Compensatory growth of fast-growing willow (Salix) coppice in response to simulated large herbivore browsing. Oikos 113: 33-42.
- Massad TJ (2013) Ontogenetic differences of herbivory on woody and herbaceous plants: A meta-analysis demonstrating unique effects of herbivory on the young and the old, the slow and the fast. Oecologia 172: 1-10.
- Averett JP, Endress BA, Rowland MM, Naylor BJ, Wisdom MJ (2017) Wild ungulate herbivory suppresses deciduous woody plant establishment following salmonid stream restoration. Forest Ecology and Management 391: 135-144.
- Flaherty KL, Rentch JS, Grafton WN, Anderson JT (2018) Timing of white-tailed deer browsing affects wetland plant communities. Plant Ecol 219: 313-324.
- Rea RV, Gillingham MP (2001) The impact of the timing of brush management on the nutritional value of woody browse for moose alcesalces. Journal of Applied Ecology 38: 710-719.
- Hanley ME, Fegan EL (2007) Timing of cotyledon damage affects growth and flowering in mature plants. Plant Cell Environ 30: 812-819.
- Meiman PJ, Thorne MS, Skinner QD, Smith MA, Dodd JL (2009) Wild ungulate herbivory of willow on two national forest allotments in wyoming. Rangeland Ecology & Management 62: 460-469.
- De Jager NR, Cogger BJ, Thomsen MA (2013) Interactive effects of flooding and deer (Odocoileus virginianus) browsing on floodplain forest recruitment. Forest Ecology and Management 303: 11-19.
- Kaufman JB (2002) Research/evaluation restoration of NE oregon streams: Effects of livestock exclosures (corridor fencing) on riparian vegetation, stream geomorphic features, and fish populations. University of North Texas, Denton, Texas, USA.
- Holland KA, Leininger WC, Trlica MJ (2005) Grazing history affects willow communities in a montane riparian ecosystem. Rangeland Ecol Manage 58: 148-154.
- Opperman JJ, Merenlender AM (2001) Deer herbivory as an ecological constraint to restoration of degraded riparian corridors. Restoration Ecology 8: 41-47.
- Hall J, Pollock M, Hoh S (2011) Methods for successful establishment of cottonwood and willow along an incised stream in semiarid Eastern Oregon, USA. Ecological Restoration 29: 261-269.
- Herder MD, Virtanen R, Roininen H (2004) Effects of reindeer browsing on tundra willow and its associated insect herbivores. Journal of Applied Ecology 41: 870-879.
Citation: Markley J, Piontek B, Coffman G (2019) Herbivory and Soil Properties as Ecological Constraints to Willow Growth in a Restored Riparian Ecosystem in Coastal California. J Environ Sci Curr Res 2: 003.
Copyright: © 2019 Jacqueline Markley, et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.