Hippocampal Microenvironment Induces the Differentiation and Polarization of Adhesive Cells between Wild Type and reeler Mice
*Corresponding Author(s):
Jinbo DengNational Health Commission Key Laboratory Of Birth Defect Prevention, Henan Key Laboratory Of Population Defects Prevention, Scientific And Technical Institute Of Population And Family Planning, Zhengzhou, China
Tel:+86 37186029589,
Fax:+86 37165961650
Email:jinbo_deng@163.com
Abstract
To investigate the effects of hippocampal microenvironment, especially Reelin on cell differentiation and cell polarization, mouse (wild type and reeler) hippocampal slices were co-cultured with various kinds of cells with different degrees of differentiation, such as the stem cells (undifferentiated cells), tumor cells (poorly differentiated cells) and terminal differentiated cells (mature somatic cells). We found that the co-cultured cells in WT group could adhere and distribute on hippocampal slices with lamination, that is, they densely adhered along pyramidal layer and granular layer, but dispersed loosely elsewhere. On the other hand, the cells in control group (cultured on blank Millipore membrane) were distributed densely and evenly, as compared to WT group and reeler group (P<0.01). In addition, compared with control and reeler group, co-cultured cells usually could differentiate and polarize under the microenvironment of WT hippocampal slices. Compared with WT hippocampal slices, the lamination of adhesive cells on reeler hippocampal slices was not obvious, and the adhesive cells were mostly distributed evenly, either for undifferentiated stem cells, poorly differentiated tumor cells or well differentiated terminal mature cells. After co-culturing for a while, the neuronal-like cells also appeared on reeler slice for iPSCs and BMSCs, but their number decreased greatly (P<0.01), and cells’ orientation was in disorder. Our data suggest that hippocampal microenvironment can induce cell differentiation and polarization, especially for the undifferentiated cells and poorly differentiated cells, and the cell differentiation and polarization are affected by both microenvironment and the degree of cell differentiation. On the other hand, the cell differentiation and polarity on reeler slice are delayed greatly with irregular orientation, indicating that Reelin promote cell differentiation and polarity.
Keywords
Cell differentiation; Cell polarization; Cell-slices co-culture; Hippocampal microenvironment; Reelin
INTRODUCTION
Cell differentiation and polarization are very important for us to understand some crucial biological and medical issues, such as ontogeny, diseases and cancer generation. In fact, cell differentiation is the processes by which cell-specific proteins, specific morphology, and cell functions are generated during the development. Cell polarization is an important property of cell differentiation. These differentiated cells can produce specific physiological or morphological characteristics. For instance, the sub-cellular structures or molecules in the polarized cells distribute asymmetrically [1,2]. Initially, differentiated cells display polarity and perform specific functions. A variety of genes are known to be involved in cell differentiation and polarity, such as Notch gene, Wnt gene, Hes gene, PAR gene, etc. [3-6]. The proteins involved in regulating cell polarization are called polarity proteins, among which the PAR family is well known [5,7]. However, as a stop signal for neural migration, Reelin is involved in cell polarity in neocortex and hippocampus [8,9]. For instance, when Reelin gene is lost in reeler mice, their main phenotype is cellular disorder in cortical plate in neocortex and hippocampus [10]. The migrating neurons cannot cross cortical plate, causing a large amount of neurons to accumulate in the subplate to form cortical inversion [11]. The phenomena above indicate that Reelin may be involved in the neuronal polarity as a signaling molecule in the microenvironment of brain. Unfortunately, Reelin’s function for cell differentiation and polarization has not gotten significant attention.
In previous studies, cell differentiation and polarization are mainly studied with cell culture, molecular biological methods, and embryological assays. However, organotypic slice culture has its own advantages as compared to conventional cell culture. For instance, it can maintain the organ’s histological structure and keep the tissue’s microenvironment as well [12,13]. With cell slice co-culture, the previous study has shown that the adhesive cells could be differentiated into neuron-like cells under induction of hippocampal microenvironment [14]. In order to further understand Reelin’s function to regulate cell differentiation and polarity, various adhesive cells were co-cultured with hippocampal slice of wild type mice or reeler mice. Therefore, the cellular differentiation and polarization under WT or reeler hippocampal microenvironment could be observed. Meanwhile, we compared the differentiation capacity of various cells, such as iPS cell (induced Pluripotent Stem Cell), Bone Marrow Mesenchymal Stem Cell (BMMSC), PC12 cell (Pheochromocytoma Cell), liver cancer 7721 cell, Primary Nervous Cell (PNC) and Primary Liver Cell (PLC). The results will provide us with new insight to understand not only the relationship between microenvironment and cell differentiation, but also the function of Reelin on cell differentiation and cell polarity.
MATERIALS AND METHODS
Animals and hippocampal slices preparation
Animals
C57BL/6J mice were used for organotypic slice preparation. The mice were bred in experimental animal center of the institute. Male and female mice were placed in breeding cages in a standard laboratory animal housing environment with a 12-h light/12-h dark cycle. All experiments were carried out in accordance with the Guidance Suggestions for the Care and Use of Laboratory Animals by Ministry of Science and Technology of the People’s Republic of China, to ensure animal welfare during experiments. Reeler mice were mutated spontaneously from C57BL/6 mice in which reelin gene were lost (Jackson Laboratory, Bar Harbor, ME, USA). The main phenotype of the mice is their unstable gait and cortical plate disorders in neocortex and hippocampus. In the study, the breeding colonies of reeler mice were produced from heterozygotes. The day of the birth of pup was defined as P0 (postnatal day 0). The genotype of reeler mutants was confirmed by PCR analysis of genomic DNA [15]. With PCR analysis, mouse tails at P5 were used for DNA samples. After tissue digestion and DNA extraction, DNA amplification was carried out, using 20 μl standard PCR identification system. The primers were 5'-CAG TTG ACA TAC CTT AAT-3' (For-wt), 5'-TGC ATT AAT GTG CAG TGT-3' (Rev-reeler), and 5'-TAA TCT GTC CTC ACT CTG CC-3' (Rev-wt). Genotype results of electrophoresis were either wild type (reelin+/+) with a 265 bp band, heterozygous (reelin+/-) with 265 bp and 363 bp bands, or homozygous (reelin-/-) with a 363 bp band.
Hippocampal slices preparation
The pups of either wild type (reelin +/+) or homozygous (reelin -/-) at P7 (postnatal day 7) were scarified for slice preparation. Initially, coronal hippocampal slices (350 µm) were prepared with a Mcllwain Tissue Chopper. Then, the hippocampal slices were transferred onto Millipore membranes insert (0.4 μm), which were placed in a six-well plate with 1 ml nutrition medium (25% horse serum, 25% Hank’s balanced salt solution, 50% minimal essential medium, 2 mmol/l glutamine, pH 7.2). The slices were cultured with 5% CO2 at 37°C for further cell-slice co-culture.
Cell preparation
According to experimental needs, three kinds of cells were used for cell adhesion analyses, the stem cells (undifferentiated cells), tumor cells (poorly differentiated cells) and terminal differentiated cells (mature somatic cells). The stem cells included iPS cell and BMMS cell; the tumor cells included PC12 cell and liver cancer 7721 cell; the mature terminal cells included primary nervous cell and primary liver cell.
7721 Cell and PC12 cell preparation
Two kinds of tumor cells were chosen to be co-cultured with hippocampal slices. Liver cancer 7721 cell is fully named as human Hepatocellular Carcinoma (HCC) cell line SMMC-7721 (SMMC-7721) which is a cell line of primary liver cancer with poor differentiation and strong division. In this study, we have chosen SMMC-7721 cell line as a target to study the cell differentiation under hippocampal microenvironment. On the other hand, rat adrenal Pheochromocytoma cell (PC12 cell) which has been generally acknowledged as nervous cell model was also used in this study. After taking 7721 cells and PC12 cells from liquid nitrogen, these cells were thawed and recovered quickly. Then, the cells were centrifuged at 1000r/min for five minutes. After supernatant was discarded, the cells were cultured in culture flasks under 5% CO2 at 37°C. Medium (pH 7.2 -7.3) was made up with 90% high glucose DMEM (GIBCO, 823186) and 10% fetal bovine serum (Beijing Soledad Technology Co., Solarbio, 111023), and the medium was changed every 2-3 days. After reaching a certain density, the cells were sub-cultured timely for subsequent cell staining (green) and cell slice co-culture.
Preparation of primary liver cells and primary nervous cells
As mature somatic cells, liver cells and nervous cells were prepared for co-culture with hippocampal slices. After liver and neocortex (P4, WT mice) were removed and washed with PBS, a piece of liver or cortex tissue was cut into small pieces with ophthalmic scissors. Then, the liver or cortical tissues were digested into disassociated cells, with 0.25% trypsin (5-fold volume of tissue) at 37°C for 10 minutes. After supernatant was discarded, the precipitated cells were washed with DMEM to fully terminate the enzyme activity. Then, suspended cells were seeded and cultured in poly-lysine coated flasks with medium (liver cells: 90% DMEM + 10% FBS; nervous cells: 80% DMEM + 10% FBS + 10% horse serum). The culture medium and non-adherent cells were removed after 24 hours. After fresh culture medium was added, the adherent cells were continuously cultured, and the medium was changed every 2 days [16]. The cultured cells were waiting for further cell-slice co-culture.
Bone marrow mesenchymal stem cell preparation
P40 mice were used for Bone Marrow Mesenchymal Stem Cell (BMMSC) preparation. After disinfection with 75% alcohol, the femur and tibia were ticked and washed repeatedly with PBS. After exposing the bone marrow cavity, a needle was penetrated inside the cavity repeatedly, and the bone marrow was washed out with L-DMEM medium containing 10% FBS. Then, the bone marrow was centrifuged at 1000 r/min for five minutes. After the supernatant was removed, the precipitated cells were pipetted repeatedly with L-DMEM medium. The suspended cells were seeded and cultured in culture flasks with 5% CO2. After 24 hours, the cultured cells without adhesion were discarded. Then, the cells were passed for two generations; large parts of bone marrow mesenchymal stem cells were produced for cell tracker CMFDA (green) staining and cell-slice co-culture.
In the meantime, in order to identify BMMSC with immunocytochemistry, the dense cells were smeared evenly on polylysine-treated glass. Then, the cells were fixed with 4% paraformaldehyde for two hours. Mouse monoclonal antibody anti-SOX2 (1:200, Abcam, AB92494) was added and incubated with cells at 4°C overnight. Next, goat anti-mouse IgG conjugated Alexa 568 (1:600, Invitrogen, AI1004) was incubated at room temperature for three hours. After rinsing three times with PBS, the slides were mounted in 65% glycerol containing DAPI (1:10000; 4' 6-diamidino-2-phenylindole). Therefore, all the cytoplasm of cells was stained with cell tracker CMFDA (green) (in detail, see also cell labeling), and the nuclei of cells could be counterstained with DAPI. Only BMMSCs could be double labeled with SOX (red) and cell tracker CMFDA (green). Under ultraviolet, fluorescein isothiocyanate (FITC) and rhodamine excitation, BMMSC could be recognized with Olympus fluorescence microscope (BX61, Japan) (Figure 1 A-F). More than 98% cells belonged to marrow mesenchymal stem cells.
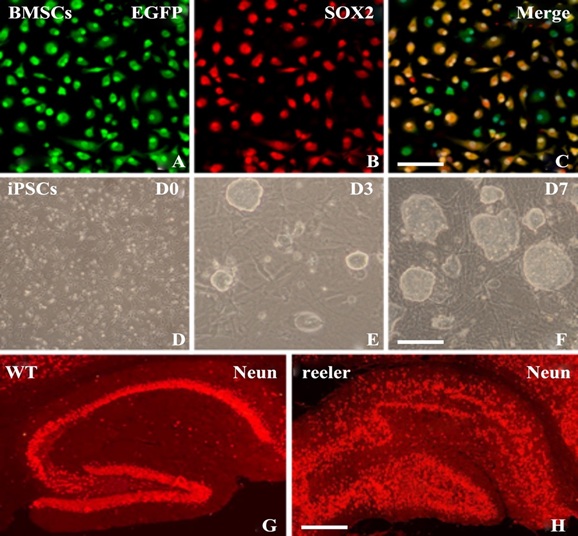 Figure 1: Stem cells and hippocampal structure.
Figure 1: Stem cells and hippocampal structure.
A-C: Identification of cultured BMSCs (tracker green and immunolabeling). All cells in photos were stained with cell tracker green CMFDA (green, A), but only BMSCs could be visualized with Sox2 immunolabeling (red, B). Photo C was merged with A and B; most cells were double stained with cell tracker green and Sox2 (C). D-F: iPS cells were cultured at various days after plating onto MEF feeder cells. D0, D3 and D7 represent cell culture at day 0, day 3 and day 7 respectively. Many Embryoid Body (EB) could be found after day 3. G & H: hippocampal structure of WT and reeler mice (NeuN immunostaining with sagittal section at P20): The pyramidal layer and granule layer (red) in WT hippocampus were hooked to each other with an obvious double “C” like shape (G). While in reeler mice, since the Reelin gene is lost, cells were arranged in disorder with no obvious formation of cell layer (H). Scale bar= 50μm in A-C, 25 μm in D-F, 250μm in G-H.
iPSC preparation
Mouse iPSCs were gifted from Dr. & Prof. D Pei (Guangzhou Institutes of Biomedicine and Health, Chinese Academy of Sciences). Mouse Embryonic Fibroblasts (MEFs) were used as feeder from C57BL/6 mice at embryonic 14 day (E14) as described previously [17]. At first, MEFs were grown in 60 mm dish for one day before iPS cells culture, and then iPS cells were seeded on the feeder cells and cultured with MEFs for 5-6 days. iPS cells medium was prepared with following formula: Knockout DMEM (Gbico, 108290180) supplemented with 20% Knockout Serum Replacement (KSR; Gbico, N10828028), 1% GlutaMAX (Gbico, 35050-061), 1% nonessential amino acids (Gbico, 11140-050), 0.1 mM 2-mercaptoethanol (Sigma, M3148), LIF recombinant mouse protein (Gbico, PMC9484). Once iPS cell clones were covered with MEF feeder (usually at day 6-8), the cells were digested into disassociated cells with 0.25% trypsin-EDTA (Gibico, 25200-056). Then, iPS cells and feeder cells suspension was transferred to dish. After differential adhesion twice (30 min for each), the feeder cells were removed, and pure iPS cells were obtained. Finally, iPS cells were resuspended in 10 ml EBs differentiation medium, then 300 μl iPS cells were plated in 96-well plate with low cell-adhesion (3000 cells/well) and were incubated at 37°C, under 5% CO2. EBs differentiation medium was prepared as following formula: Knockout DMEM, 20% Knockout serum replacement, 1% GlutaMAX, 1% Nonessential amino acids, 0.1 mM 2-mercaptoethanol, bFGF (4 ng/ml) (Gbico, PMG0034). Fresh medium was changed every two days. After several days, cultured iPSCs became dense, and some cells gathered together to form Embryoid Body (EB), and the iPSCs were ready for continuous experiments, cell staining with cell tracker green CMFDA and cell-hippocampus co-culture.
Cell labeling and cell-slice co-culture
Cell labeling
The experimental cells were iPSCs, BMSCs, PC12 cells, SMMC-7721 cells, primary neural cells, and primary liver cells. Cell Tracker fluorescent probes (1:1000, Life Technologies Corporation, C2925) can be used to stain cells with green color. The following was the details for cell staining. Initially, the cells for co-culture were digested with 0.25% trypsin respectively. Then, the cells were pipetted and rinsed for one to two minutes with serum-free DMEM medium and labeled with cell tracker green CMFDA at final concentration of 5 μM. The cells were incubated at 37°C for 20 minutes in the dark place and rinsed with DMEM containing 10% FBS. The cells were centrifuged three times at 1000 r/min and dispersed by pipetting. Finally, the cell concentration with 1×106 cells /ml cells was prepared for cell-slice co-culture.
Cell-slices co-culture
In order to understand the effects of hippocampal microenvironment with or without Reelin on cell differentiation and polarity, different cells were co-cultured with WT hippocampal slices or reeler slices. Slice culture was described as above. After a few hours of slice culture, the cell suspension stained with Cell Tracker green was dropped onto the hippocampal slices. After incubation for 15 minutes, slices were washed three times with culture medium. Finally, the cell-slice was continuously co-cultured with 5% CO2 at 37°C for another six hours to several days. Under hippocampal microenvironment, the cell differentiation would be observed in iPSCs, BMMSCs, 7721 cells, PC12 cells, primary liver cells, and primary nervous cells on slices. On the other hand, the cells mentioned above were cultured on Millipore membrane directly as control. Therefore, experiments were divided into three groups, WT group (adherent cells were cultured on hippocampal slice of WT mouse), reeler group (adherent cells were cultured on hippocampal slice of reeler mouse) and control group (cells were cultured on blank Millipore membrane). After experiments, the cultured slices with Millipore membrane were rinsed three times with PBS and fixed in 4% paraformaldehyde at 4°C overnight in the dark. Finally, co-cultured slices were mounted with 65% glycerol plus DAPI (1:5000) for imaging. In this way, the hippocampal structure could be counterstained with DAPI. The images were made with fluorescent microscope (Olympus, BX61, Japan) by FITC, rhodamine and ultraviolet excitation. The cells in control groups were also prepared as above.
Measurements and statistical analyses
After taking images under microscope, measurements were carried out with ImageJ software. The following parameters were used to evaluate the degree of cell differentiation and polarity. At least 10 slices were used for measurement in each group. 1) Cell density was used to evaluate the cell number after co-culture. Cell density = cell number/area (cells/mm2). 2) Rate of polarization was used to quantify the cell polarization. Rate of polarization = number of polarized cells/total cell number in same area. 3) On the other hand, the degree of cell polarization was measured with the variation of polarized cell (PV). PV= length/cell width.
In order to understand the differentiation of cells under hippocampal microenvironment with or without Reelin, the parameters mentioned above were compared among various cells, such as iPSC BMMSC, 7721 cell, PC12 cell, PNC and PLC. The data were recorded in a Microsoft Excel spreadsheet. Inter-group comparisons among three groups (control group, WT group and reeler group) were performed using the one-way ANVOA q test with SPSS 17.0 statistical software. P
RESULTS
Distribution and lamination of adherent cells on WT or reeler hippocampal slices
The hippocampus is an archicortex with three-layer lamination. For coronal section, hippocampus consists of double C shapes which hooked to each other forming Ammon's horn and dentate gyrus (Figure 1G). After co-culture, the cell density on WT hippocampal slices, either for undifferentiated stem cells (such as iPSCs and BMMSCs), poorly differentiated cells (such as liver cancer SMMC-7721 cells, PC12 cells), or terminal well differentiated cells (such as PNCs and PLCs), were significantly lower as compared to the control group which grew without cell-slice contact inhibition (Figure 2A, Figure 3A). Furthermore, the co-cultured cells on WT hippocampal slices demonstrated orderly distribution. The adherent cells in WT group appeared as similar hippocampal lamination, that is, the cells on the pyramidal layer or granular layer were much denser than elsewhere (Figure 2B, Figure 3B). The adherent cells in control group which were cultured on the blank Millipore membrane were very dense and distributed without lamination (Figure 2A, Figure 3A). On the other hand, the various cells were also co-cultured with reeler hippocampal slices. Lacking Reelin protein in the hippocampus, the hippocampal structure changed greatly. For instance, the hippocampal lamination was in disarray, especially in the dentate gyrus. Granule cells dispersed toward both the molecular layer and hilus, and the double C shape, which consisted of the granule and pyramidal layers, was replaced by a drumstick-like structure (Figure 1H) [18]. In our study, compared to the WT group, the lamination of adherent cells on reeler slices was not obvious, and the cells were mostly distributed evenly on slices, either for undifferentiated stem cells, poorly differentiated tumor cells or differentiated terminal cells (Figure 2C, Figure 3C).
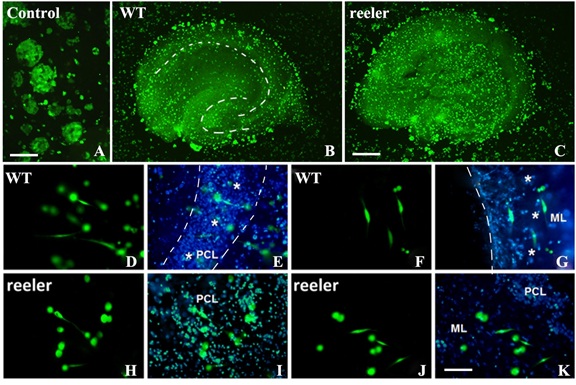 Figure 2: Lamination and polarization of iPS cells under induction of hippocampal microenvironment.
Figure 2: Lamination and polarization of iPS cells under induction of hippocampal microenvironment.
A-C: Cell co-culture on WT (B) and reeler (C) hippocampal slices or without slices (control group) (A). In control group, iPSCs were cultured on Millicell membrane directly, after culture for one day, the round cells scattered on membrane, and some cells aggregated together to form embryoid bodies (A). When iPSCs were co-cultured for 24 hours with WT hippocampal slice (B), the adhesive iPSCs on slice showed hippocampal lamination with double “C” shape, and the cells concentrated mainly on granular cell layer and pyramidal layer with high density. When iPSCs were co-cultured with reeler hippocampal slices, the lamination of cells was lost. D-G: With high magnification, the differentiation and polarization of iPS cells under induction of WT hippocampal slice. After co-culture for 24 hours, iPSCs started to differentiate into neuron-like cells with obvious polarization. For instance, in pyramidal cell layer (PCL is located between two dotted lines and marked with * as well), the differentiated neuron-like cells were oriented vertically with similar shape of pyramidal cell in vivo (D & E). However, cells on molecule layer on right side of dotted line (ML, marked with * as well) were arranged horizontally with fusiform shape (F & G). H-K: the differentiation and polarization of iPS cells under induction of reeler hippocampal slice. Under induction of reeler hippocampal microenvironment, the differentiations of iPS cells also occur, but their polarization is delayed, and the orientation is in disorder on either pyramidal cell layer or molecular layer (H-K). H & I show the differentiation of iPS cells on pyramidal cell layer (PCL), and J-K show the differentiation of iPS cells on molecular layer (ML). Bar=50 μm in A, 250 μm in B & C; 50μm in D-K.
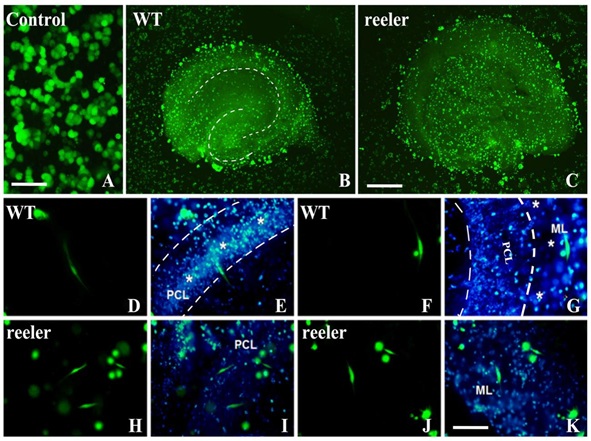 Figure 3: The lamination and polarization of BMMS cells under induction of hippocampal microenvironment.
Figure 3: The lamination and polarization of BMMS cells under induction of hippocampal microenvironment.
A-C: Cells were co-cultured with WT (B) and reeler (C) hippocampal slices or without slices (control group) (A). In control group, where BMMSCs were cultured on Millicell membrane directly, after culture for one day, cells kept round or polygonal shapes (A). When BMMSCs were co-cultured for 24 hours with WT hippocampal slice (B), the adhesive cells showed hippocampal lamination with double “C” shape, and the cells were concentrated mainly on granular cell layer and pyramidal layer with high density. When BMMSCs were co-cultured with reeler mice, the lamination of cells was lost (C). D-G: The differentiation and polarization of BMMS cells under induction of WT hippocampal slice (hippocampal structure was counterstained with Dapi, blue color). After co-culturing for 24 hours, BMMSCs started to differentiate into neuron-like cells with obvious polarization. For instance, in pyramidal cell layer (PCL, between two dotted lines and marked with * as well), the differentiated neuron-like cells were oriented vertically with similar shape of pyramidal cell in vivo (D & E). E is the same photo of D plus Dapi counterstaining. However, cells on molecule layer (ML, on the right side of PCL and marked with * as well) were arranged horizontally (F & G). G is the same photo of F plus Dapi counterstaining. H-K: The differentiation and polarization of BMMS cells under induction of reeler hippocampal slice (cell tracker green CMFDA and Dapi counterstaining). Under induction of hippocampal microenvironment of reeler mice, the differentiation of BMMS cells could occur as in WT mouse, but their orientation was in disorder on either pyramidal cell layer or molecular layer (H-K), and their processes and rate of polarization were less than WT group. The differentiation of BMMS cells on pyramidal layer (PCL) of reeler slice is shown in H & I, and so did BMMS cells in molecular layer in J & K. Bar=50 μm in A; 350 μm in B & C; 50 μm in D-K.
Cell density was quantified statistically with Image J software. The cell density was measured among various kinds of cells, and statistical tests were compared between WT group and control group. The cell density in the control group was much higher than the cells on hippocampal slices (P<0.01), suggesting that hippocampal tissue inhibited the proliferation of adherent cells (Figure 4). On hippocampal slices, the cell density of iPSCs, BMMSCs, SMMC-7721 cells and PC12 cells was higher than primary liver cells and primary neural cells (Figure 4) (P<0.05). This suggests that poorly differentiated cells had high affinity to hippocampal slices and had high proliferation.
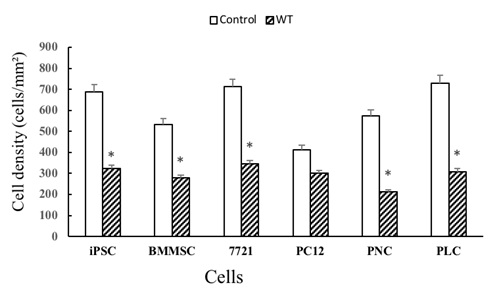 Figure 4: Cell densities in various groups.
Figure 4: Cell densities in various groups.
Cell densities on membranes (control groups) were higher than the cells on WT hippocampal slices (P< 0.01), suggesting hippocampal microenvironment had inhibitory effect for cell proliferation. iPSC: Induced pluripotent stem cell; BMMSC: Bone marrow mesenchymal stem cell; 7721: liver cancer 7721 cell. PC12: Pheochromocytoma cell; PNC: Primary nervous cell; PLC: Primary liver cell. *: Compared with control group, P< 0.01.
Differentiation and polarity of adherent cells on WT or reeler hippocampal slices
After cell co-culture on WT hippocampal slices for a day, even hours, the stem cells and tumor cells on the slices started to differentiate and grow out some processes, at first one or two, then several. Since these cells with processes had polarization and were similar to nervous cell, we called them neuron-like cells. In high magnification, the stem cells and tumor cells could differentiate into neuron-like cells with obvious polarization (Figure 2 D-G, Figure 3 D-G). Interestingly, their arrangement and polarization were not in disorder, but showed clear orientation. For instance, the adherent cells on molecular layer were often arranged horizontally and looked like fusiform with two processes. While the cells located on the granular cell layer and pyramidal cell layer were arranged vertically and looked like triangles with multiple processes, as the neurons in vivo (Figure 5 A-N). The poorly differentiated tumor cells were similar to the stem cells, but their degrees of differentiation and polarity were a little lower than stem cells (Figure 5 A-N). However, compared with stem cells and tumor cells, well differentiated terminal cells, such as primary liver cells, seemed not to differentiate and polarize, and most of cells kept their spherical shape without processes (Figure 5 R-T). The results suggested that the differentiated terminal liver cells had weak capacity for differentiation. Unlike primary liver cells, primary nervous cells seemed to be an exception. As differentiated terminal cells, primary neural cells could produce polarization as stem cells and tumor cells, since these cells were themselves neurons which could restore their original shape under hippocampal microenvironment (Figure 5 O-Q). On the contrary, in the control group where adherent cells were cultured on Millipore membrane, the cell differentiation and polarization seemed not to happen, even for stem cells and tumor cells (Figure 4 A,D,G,L,Q,R). The rate of polarization in various cells was measured and calculated. The data showed that under induction of WT hippocampal microenvironment, the stem cells, tumor cells and even PNCs had high rates of polarization, due to their high ability for cell differentiation (Figure 6 A,B) (P<0.01). Furthermore, the degree of cell differentiation was also evaluated with variation of polarization. Under the induction of WT hippocampal microenvironment, stem cells, tumor cells and PNCs showed higher variation of polarization and larger size than PLCs (Figure 4 A-N, Figure 6B) (P<0.01), suggesting that they had strong differentiation ability.
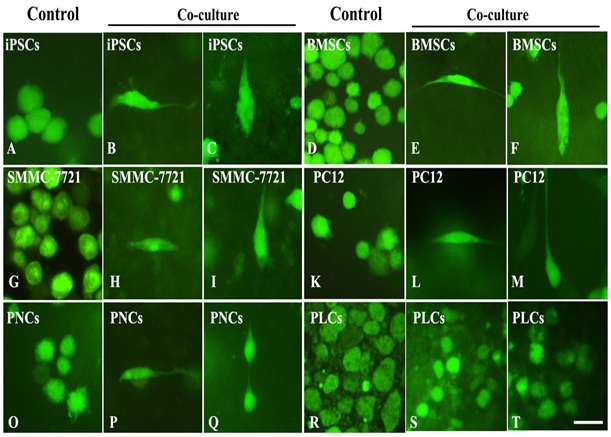 Figure 5: Distribution and differentiation of co-cultured cells on WT hippocampal slices with high magnification.
Figure 5: Distribution and differentiation of co-cultured cells on WT hippocampal slices with high magnification.
A-C: the differentiation and polarization of iPSCs’. In control group (A), the cells appeared round without polarization, but under WT hippocampal microenvironment, iPSCs could differentiate into neuron-like cells. For instance, the cells could polarize into neuron-like cells with horizontal orientation (B) on molecular layer, or vertical arrangements (C) on pyramidal layer. D-F: the differentiation and polarization of BMMSCs’. In control group (D), as iPSCs, BMMSCs appeared round without polarization. Under induction of WT hippocampal microenvironment, BMMSCs can also produce polarization on molecular layer with horizontal orientation (E) and on pyramidal layer with vertical arrangement (F). G-I: the differentiation and polarization SMMC-7721 cells. The cells in control group looked round without differentiation (G). However, under induction of WT hippocampal slice, some cells on molecular layer polarized into neuron-like cell with fusiform shape (H) and triangle shape (I) on pyramidal layer. L-N: the differentiation and polarization of PC12 cells. L: No-polarized cells with round shape in control group. M: Polarized cells on molecular layer. N: Polarized cell on pyramidal layer. O-Q: the differentiation and polarization of primary nervous cells. Photo O shows the no-polarized cells in control group. Even though the primary nervous cells belonged to terminal cells, under induction of hippocampal slices, these primary nervous cells could produce cell polarization with horizontal orientation (P) on molecular layer and vertical arrangement (Q) on pyramidal layer. R-T: the differentiation and polarization of primary liver cells. Since the primary liver cells were terminal differentiated cells, they almost lost the capacity of differentiation and polarization. Therefore, the cells with round shape in either control group (R) or WT group (S & T). S: The cells were on molecular layer. T: The cells were on pyramidal layer. Bar = 30 μm in A-T.
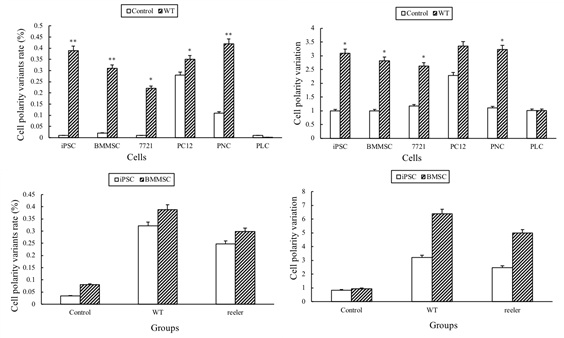 Figure 6: Rate of polarization and variation of polarization among various groups
Figure 6: Rate of polarization and variation of polarization among various groups
A: The rate of polarization for iPSC, BMMSC, 7721 cell, PC12 cell, PNC and PLC. The cellular rates of polarization in WT groups were higher than control groups (P<0.01), suggesting hippocampal microenvironment can induce cell differentiation and polarization. Furthermore, compared with terminal differentiated cell (for example, primary liver cell), the rates of palarization for stem cells and tumor cells were higher than primary liver cell (P< 0.01), suggesting that stem cells and tumor cells have strong capacity for cell polarization. B: Cells’ variation of polarization between control group and WT group. As rate of polarization, the cellular variation of polarization in WT group is higher than control (comparison in same kind of cell) (P<0.01), but among different kinds of cells, stem cells and tumor cells have higher variation of polarization than PLC (P<0.01). C: the rate of polarization of iPSCs and BMSCs’ among control group, WT group and reeler group. iPSCs and BMSCs in reeler group have higher rate of polarization than control group, but lower than WT group (P<0.05), suggesting that microenvironment without Reelin delayed cell differentiation and polarization. D: the variation of polarization of iPSCs and BMSCs’ among control, WT group and reeler group. Like rate of polarization, iPSCs and BMSCs in reeler group have higher variation of polarization than control group, but lower than WT group (P<0.05), suggesting that microenvironment without Reelin delayed cell differentiation and polarization.
In order to understand the effect of Reelin on cell differentiation and polarity, we examined their difference in cell polarization among reeler slice, WT slice, and control respectively. In this study, various adherent cells were co-cultured with reeler hippocampal slices. After co-culture for one day, the neuronal-like cells also appeared on reeler slice for iPSCs, BMMSCs, PC12 cell, liver cancer 7721-cells and PNCs. But unlike in WT group, the number of neuronal-like cells decreased, and the orientation of cells was not regular (Figure 2 H-K, Figure 3 H-K). The adherent cells on granular layer and the pyramidal cell layer usually showed non-vertical arrangement (Figure 2 H-K, Figure 3 H-K), and the cells on molecular layer were also not arranged horizontally (Figure 2 H-K, Figure 3 H-K). In order to understand the effects of Reelin on cell polarity, we examined the rate of polarization and variation of polarization of cells in reeler group and compared them with WT and control groups. Both the rate of polarization and variation of polarization in reeler groups were higher than those in control group, (Figure 6 C,D) (P<0.01), suggesting that cell differentiation and polarity could be induced under hippocampal microenvironment even without Reelin. However, in WT groups, the rate of polarization and variation of polarization were higher than reeler group (P<0.05), indicating that the hippocampal microenvironment with Reelin could accelerate cell polarization.
DISCUSSION
Cell differentiation and cell polarization are hot topics in cell biology and neuroscience. Cellular differentiation and polarization are inseparable, and cell polarization is regarded as a morphological phenotype of cell differentiation. Cell polarization is a very popular biological phenomenon. For instance, the columnar epithelial cells in small intestinal mucosa can be recognized with a clear top (apical side) and bottom (basal side) [19,20], and a neuron has an axon and multiple dendrites as well. In fact, the cell polarization can play important roles in cellular physiology, such as determining the direction of cell migration and electric impulse from dendrites to axon. As we know, stem cells have strong capacity for cell differentiation and cell polarity. For example, fertilized eggs have the totipotent potential for differentiation, and progenitor cells have a tendency to differentiate into specific tissues and cells [20]. Cellular polarization and differentiation are regulated by some proteins. These proteins are called polarity proteins. PAR (Partitioning defective) family is typical and studied well, which includes PAR-3, PAR-6, and atypical protein kinase C (atypical protein kinase C, aPKC) [21]. PAR functions through PAR-3/PAR-6/PKC-3 pathways [22]. According to previous studies, aPKCis widely present in nervous tissue and expressed mainly in axon, dendrite, and synapse, suggesting that PAR protein plays an important role in establishing neuronal polarity [23,24]. Recently, Reelin is believed as a potential polarity molecule for neuronal differentiation and polarization [25], but more evidence is needed to support the hypothesis. Cell culture is often used to investigate cell differentiation and cell polarization. Unfortunately, the cellular model hardly mimics the real situations during cell differentiation, such as histological structure and tissue microenvironment. With cell-slice co-culture, the hippocampal slices provide a properly experimental model for the study of cell differentiation and polarization. In the study, various kinds of cells were co-cultured with either non-slice, WT hippocampal slices or reeler hippocampal slices respectively. Therefore, the cell polarization and differentiation could be induced, and the cell differentiation and cell polarization could be investigated well under the effects of various cell types and different microenvironments.
Hippocampal microenvironment with Reelin induces the adhesive cells to form lamination
In our study, compared with WT control, the co-cultured adhesive cells were distributed on hippocampal slices with lamination; that is, they were densely adhered along pyramidal layer and granular layer but loosely elsewhere. However, in the control group, the cells were distributed densely and evenly with round shape, indicating that there was proliferation inhibition for co-culture cells on hippocampal slices. The laminar distribution of cells was probably caused by some adhesive molecules in hippocampus, and these adhesive molecules were distributed as hippocampal lamination [26]. There are several kinds of adhesive molecules, and Neural Cell Adhesion Molecule (NCAM), cadherin, selectin, integrin, hyaladherin are important in central nervous system. These molecules have multiple functions for CNS development; for example, cell shaping, cell migration and neurite growth [27]. Previous study also showed that neuron growth in retina was clearly laminin-dependent [28], and the neurons had great affinity to laminin [28]. Besides the affinity to cells, the adhesive molecules can inhibit cell division and regulate cell differentiation, which is important for the sculpting of cells, tissues and organs during embryonic development [29]. Some other extracellular matrixes are also involved in the cell polarization, such as LKB1 [30]. It is important for neurite outgrowth and axon formation, and the overexpression of LKB1 causes multiple axons in a neuron. On contrary, Sema3A can inhibit the formation of axons by down-regulating protein kinase PKA-dependent LKB1 phosphorylation [31]. Therefore, according to the analyses above, it is reasonable that, in control group, without the contact between adhesive cells and hyppocampal slice, the cells proliferate very quickly and densely, and once adhesive cells make contact with hippocampal slice, the cell proliferation was sharply decreased. In addition, the terminal differentiated cells, such as neuronal cells and liver cells show less proliferative ability, the cell density in both control group and WT group is much lower than stem cell and poorly differentiated tumor cell.
Hippocampal microenvironment with Reelin induces stem cells and tumor cells to differentiate into neuron-like cells
Hippocampal microenvironment with Reelin can induce not only the distribution of adhesive cells with lamination, but also induce adhesive cells to differentiate and polarize. In control group, it seems that no differentiation and polarization occur in the cultured cells, and these cells always keep round shape. In WT group, after co-culture a while, the adhesive cells started to differentiate and polarize on the hippocampal slices. For instance, stem cells (iPSCs and BMMSCs) and poorly differentiated cells (SMMC-7721 cell and PC12 cell) could grow into neuron-like cells with polarity; however, well-differentiated terminal cells, such as PLCs, were not. The results suggest that stem cell and poorly differentiated tumor cell had higher capacity for cell polarization than terminal differentiated cells. Stem cells and tumor cells can be self-renewed. So, these cells can differentiate and polarize under induction of hippocampal microenvironment. Generally, terminal differentiated cells can express their functional specific proteins, and they gradually lose differentiation potential. In the study, the primary liver cells could not produce polarization on hippocampal slices; therefore, they appeared round, as the cells in control group. Unlike primary liver cells, primary nervous cells seemed an exception, and they changed their round shape into neuron-like cells with polarity, even though the neuronal projections were cut off during the process of cell dissociation. It is regarded as cell repair rather than cell differentiation. PNCs try to restore their morphology and polarity under the induction of hippocampal microenvironment. Our study suggests that differentiated status of cells is also the key factor for cell polarization, and both microenvironment and cell type together determine the cell differentiation and polarization. Some polarity protein is involved the process. The polarity proteins and their receptors, such PAR family [32] and CDC42 [33,34], play an important role in the process. PAR is a protein family encoded by conserved genes par-1 to par-6, and they are the key factor in cell polarity of nematodes, fruit flies and mammalian [24,35]. In addition, serine/threonine kinase LKB1 in mammals is homologue to PAR-4 in nematode, and it is also indispensable in the neuronal polarization and migration in vivo [36]. In mammals, Numb protein, distributed in cortex and neuroepithelium, is probably involved in the cell polarization as well [37].
Disordered cell arrangement and delayed cell polarization under hippocampal microenvironment without Reelin
Reelin is an important protein in molecular layer, and it is a stop signal for radial neuronal migration [8,9,38]. Reelin knockout mice, reeler mice, have laminar disorder in cerebral cortex and hippocampus, suggesting that Reelin is important for neuronal differentiation and polarity [39]. Cell co-culture with reeler hippocampal slice may help us to understand Reelin’s functions in cell differentiation and cell polarity. In our study, the cell differentiation of reeler group was totally different from WT group. At first, the adhesive cell on reeler hippocampal slice was distributed evenly without obvious lamination. Secondly, although some neuronal-like cells could be found, but the polarized cells were much less than in WT group, especially for iPSCs and BMSCs, suggesting that the microenvironment without Reelin can delay the polarization, that is, Reelin can promote the orderly distribution of cells. The process is probably mediated by Reelin together with some adhesion molecules. Matsunaga et al., reported that Reelin could mediate neuronal aggregation in an N-cadherin-dependent manner [40,41]. With atomic force microscopy, the authors found that Reelin indeed enhanced the adhesiveness of neurons to produce N-cadherin. As adhesion molecule, N-cadherin is densely concentrated along granular layer and pyramidal layer, to eventually attract the distribution of adhesive cells with lamination. Meanwhile, Reelin is also involved in cell polarization as well. Our study showed that in WT group, the polarized cells often were parallel to molecular layer and perpendicular to granular layer or pyramidal cell layer, but in reeler group, the orientations of cells were in disorder without definite directions. For instance, the adhesive cells on granular layer, pyramidal cell layer and molecular layer showed non-regular orientation. Quantified data show that both the rate of polarization and variation of polarization in reeler group are lower than WT group, indicating that the hippocampal microenvironment in reeler slice can delay cell differentiation and polarization. In other words, Reelin functions to promote cell differentiation and polarization. Previous studies have shown that Reelin could promote apical dendritic growth to where Cajal-Retzius cells are located [18]. Reelin probably regulates cell polarization through cytoskeletal motility and cytoskeletal reorganization [38]. Reelin can promote the formation of filopodia and neurite growth by regulating the microtubule assembly and depolymerization [42]. To carry out the works, Reelin should induces Dab1 phosphorylation through VLDLR pathway [43], and Dab1 phosphorylation further activates Rap1 which finally leads to cytoskeletal motility and cytoskeletal reorganization in neurons as well [44].
In summary, various cells were co-cultured with hippocampal slice of wild type mice or reeler mice, and the differentiation and polarization of co-cultured cells were investigated. 1) Compared to control group, the adhesive cells on WT hippocampal slices show laminar distribution as original hippocampal structure, and cell differentiation and polarization are induced as well. The stem cells and poorly differentiated tumor cells can differentiate into neuron-like cells, but terminal differentiated cells are difficult to achieve. 2) The adhesive cells can distribute on reeler hippocampal slice without lamination. Furthermore, under induction of reeler hippocampal microenvironment, some neuronal-like cells could be found in co-cultured cells, but their number is much less than the cells on WT slice, suggesting that cell differentiation and polarization are delayed. 3) Our study suggests that both hippocampal microenvironment and the cell type together determine the differentiation and polarization of cells. Microenvironment without Reelin can delay the cell polarization, since Reelin functions to promote orderly distribution of cells. Hopefully, our data can provide some valuable knowledge about cell differentiation and polarization and benefit the clinical practice of stem cell transplantation.
FUNDING
This study was funded by Scientific and Technical Project of Henan Science and Technology Department (192102310134), the Doctoral Research Foundation of Luohe Medical College (2019-DF-01) and the Open Project of National Health Commission Key Laboratory of Birth Defect Prevention (ZD201903).
ETHICAL APPROVAL
The use of animals in this study was approved by the Animal Ethical Committee of Scientific and Technical Institute of Population and Family Planning, and all applicable national guidelines for the care and use of animals were followed.
CONFLICT OF INTEREST
The authors declare that they have no competing interests.
AUTHOR’S CONTRIBUTION
Wenjuan Fan and Jinbo Deng conceived and designed this study. Wenjuan Fan, Yizheng Sun and Shanshan Wu collected and interpreted data. Wenjuan Fan and Jinbo Deng wrote the manuscript. Jinbo Deng was in charge of material supply and administrative support.
REFERENCES
- Masuda-Hirata M, Suzuki A, Amano Y, Yamashita K, Ide M, et al. (2009) Intracellular polarity protein PAR-1 regulates extracellular laminin assembly by regulating the dystroglycan complex. Genes Cells 14: 835-850.
- Pohl C (2017) The Midbody and its Remnant in Cell Polarization and Asymmetric Cell Division. Results Probl Cell Differ 61: 165-182.
- Schweisguth F (2015) Asymmetric cell division in the Drosophila bristle lineage: from the polarization of sensory organ precursor cells to Notch-mediated binary fate decision. Wiley Interdiscip Rev Dev Biol 4: 299-309.
- von Boehmer H (2005) Notch in lymphopoiesis and T cell polarization. Nat Immunol 6: 641-642.
- Bayraktar J, Zygmunt D, Carthew RW (2006) Par-1 kinase establishes cell polarity and functions in Notch signaling in the Drosophila embryo. J Cell Sci 119: 711-721.
- Mohammadpour H, Pourfathollah AA, Nikougoftar Zarif M, Tahoori MT (2015) Effects of DKK-3, a Wnt signaling inhibitor, on dendritic cell phenotype and T cell polarization. Immunopharmacol Immunotoxicol 37: 481-487.
- Goldstein B, Macara IG (2007) The PAR proteins: fundamental players in animal cell polarization. Dev Cell 13: 609-622.
- Zhao S, Frotscher M (2010) Go or stop? Divergent roles of Reelin in radial neuronal migration. Neuroscientist 16: 421-434.
- Frotscher M, Chai X, Bock HH, Haas CA, Forster E, et al. (2009) Role of Reelin in the development and maintenance of cortical lamination. J Neural Transm (Vienna) 116: 1451-1455.
- Zhao S, Chai X, Bock HH, Brunne B, Forster E, et al. (2006) Rescue of the reeler phenotype in the dentate gyrus by wild-type coculture is mediated by lipoprotein receptors for Reelin and Disabled 1. J Comp Neurol 495: 1-9.
- Andersen TE, Finsen B, Goffinet AM, Issinger OG, Boldyreff B (2002) A reeler mutant mouse with a new, spontaneous mutation in the reelin gene. Brain Res Mol Brain Res 105: 153-156.
- Kronander E, Michalski N, Lebrand C, Hornung JP, Schneggenburger R (2017) An organotypic slice culture to study the formation of calyx of Held synapses in-vitro. PLoS One 12: 0175964.
- Venkataramanappa S, Simon R, Britsch S (2015) Ex utero electroporation and organotypic slice culture of mouse hippocampal tissue. J Vis Exp 97.
- Forster E, Kaltschmidt C, Deng J, Cremer H, Deller T, et al. (1998) Lamina-specific cell adhesion on living slices of hippocampus. Development 125: 3399-3410.
- Deller T, Haas CA, Deissenrieder K, Del Turco D, Coulin C, et al. (2002) Laminar distribution of synaptopodin in normal and reeler mouse brain depends on the position of spine-bearing neurons. J Comp Neurol 453: 33-44.
- Zhao S, Chai X, Forster E, Frotscher M (2004) Reelin is a positional signal for the lamination of dentate granule cells. Development 131: 5117-5125.
- Baharvand H, Totonchi M, Taei A, Seifinejad A, Aghdami N, et al. (2010) Human-induced pluripotent stem cells: derivation, propagation, and freezing in serum- and feeder layer-free culture conditions. Methods Mol Biol 584: 425-443.
- Wu P, Li MS, Yu DM, Deng JB (2008) Reelin, a guidance signal for the regeneration of the entorhino-hippocampal path. Brain Res 1208: 1-7.
- Cereijido M, Contreras RG, Shoshani L (2004) Cell adhesion, polarity, and epithelia in the dawn of metazoans. Physiol Rev 84: 1229-1262.
- St Johnston D, Ahringer J (2010) Cell polarity in eggs and epithelia: parallels and diversity. Cell 141: 757-774.
- Hurov J, Piwnica-Worms H (2007) The Par-1/MARK family of protein kinases: from polarity to metabolism. Cell Cycle 6: 1966-1969.
- Tostevin F, Howard M (20089) Modeling the establishment of PAR protein polarity in the one-cell C. elegans embryo. Biophys J 95: 4512-4522.
- Chen YM, Wang QJ, Hu HS, Yu PC, Zhu J, et al. (2006) Microtubule affinity-regulating kinase 2 functions downstream of the PAR-3/PAR-6/atypical PKC complex in regulating hippocampal neuronal polarity. Proc Natl Acad Sci U S A, 103: 8534-8539.
- Hapak SM, Rothlin CV, Ghosh S (2018) PAR3-PAR6-atypical PKC polarity complex proteins in neuronal polarization. Cell Mol Life Sci 75: 2735-2761.
- Forster E (2014) Reelin, neuronal polarity and process orientation of cortical neurons. Neuroscience 269: 102-111.
- Rauvala H, Pihlaskari R, Laitinen J, Merenmies J (1989) Extracellular adhesive molecules in neurite growth. Biosci Rep 9: 1-12.
- Lochter A, Taylor J, Braunewell KH, Holm J, Schachner M (1995) Control of neuronal morphology in vitro: interplay between adhesive substrate forces and molecular instruction. J Neurosci Res 42: 145-158.
- Adler R, Jerdan J, Hewitt AT (1985) Responses of cultured neural retinal cells to substratum-bound laminin and other extracellular matrix molecules. Dev Biol 112: 100-114.
- Dzamba BJ, DeSimone DW (2018) Extracellular Matrix (ECM) and the Sculpting of Embryonic Tissues. Curr Top Dev Biol 130: 245-274.
- Asada N, Sanada K, Fukada Y (2007) LKB1 regulates neuronal migration and neuronal differentiation in the developing neocortex through centrosomal positioning. J Neurosci 27: 11769-11775.
- Wang N, Dhumale P, Chiang J, Puschel AW (2018) The Sema3A receptor Plexin-A1 suppresses supernumerary axons through Rap1 GTPases. Sci Rep 8: 15647.
- Dickinson DJ, Schwager F, Pintard L, Gotta M, Goldstein B (2017) A Single-Cell Biochemistry Approach Reveals PAR Complex Dynamics during Cell Polarization. Dev Cell 42: 416-434.
- Boscher C, Gaonac'h-Lovejoy V, Delisle C, Gratton JP (2019) Polarization and sprouting of endothelial cells by angiopoietin-1 require PAK2 and paxillin-dependent Cdc42 activation. Mol Biol Cell 30: 2227-2239.
- Goryachev AB, Leda M (2017) Cell Polarity: Spot-On Cdc42 Polarization Achieved on Demand. Curr Biol 27: 810-812.
- Lang CF, Munro E (2017) The PAR proteins: from molecular circuits to dynamic self-stabilizing cell polarity. Development 144: 3405-3416.
- Shelly M, Poo MM (2011) Role of LKB1-SAD/MARK pathway in neuronal polarization. Dev Neurobiol 71: 508-527.
- Breunig JJ, Rakic P (2011) Coordinating migratory neuron polarization by numb-ing communication. Dev Cell 20: 578-580.
- Frotscher M, Zhao S, Wang S, Chai X (2017) Reelin Signaling Inactivates Cofilin to Stabilize the Cytoskeleton of Migrating Cortical Neurons. Front Cell Neurosci 11: 148.
- Santana J, Marzolo MP (2017) The functions of Reelin in membrane trafficking and cytoskeletal dynamics: implications for neuronal migration, polarization and differentiation. Biochem J 474: 3137-3165.
- Matsunaga Y, Noda M, Murakawa H, Hayashi K, Nagasaka A, et al. (2017) Reelin transiently promotes N-cadherin-dependent neuronal adhesion during mouse cortical development. Proc Natl Acad Sci U S A 114: 2048-2053.
- Chai X, Forster E, Zhao S, Bock HH, Frotscher M (2009) Reelin stabilizes the actin cytoskeleton of neuronal processes by inducing n-cofilin phosphorylation at serine3. J Neurosci 29: 288-299.
- Leemhuis J, Bouche E, Frotscher M, Henle F, Hein L, et al. (2010) Reelin signals through apolipoprotein E receptor 2 and Cdc42 to increase growth cone motility and filopodia formation. J Neurosci 30: 14759-14772.
- Sharaf A, Bock HH, Spittau B, Bouche E, Krieglstein K (2013) ApoER2 and VLDLr are required for mediating reelin signalling pathway for normal migration and positioning of mesencephalic dopaminergic neurons. PLoS One 8: 71091.
- Moon UY, Park JY, Park R, Cho JY, Hughes LJ, et al. (2015) Impaired Reelin-Dab1 Signaling Contributes to Neuronal Migration Deficits of Tuberous Sclerosis Complex. Cell Rep 12: 965-978.
Citation: Fan W, Sun Y, Wu S, Fu S, Deng J (2020) Hippocampal Microenvironment Induces the Differentiation and Polarization of Adhesive Cells between Wild Type and reeler Mice. J Stem Cell Res Dev Ther 6: 029.
Copyright: © 2020 Wenjuan Fan, et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.