Herald Scholarly Open Access is a leading, international publishing house in the fields of
Science. Our mission is to provide an access to knowledge globally.
We provide highquality
articles to the scientific community and we strive for your research improvement and
distinguishment throughout the world.


Welcome to Herald Scholarly Open Access
Herald Scholarly Open Access gladly welcome's your submission
Featured Journals
Herald Scholarly Open Access is successfully running 73+ peer-reviewed open access journals in the fields of Clinical, Medical, Chemistry and Pharmaceuticals and also published 1500+ articles indexed in various sites and also cited by various authors around the world. View All Journals
Latest Articles
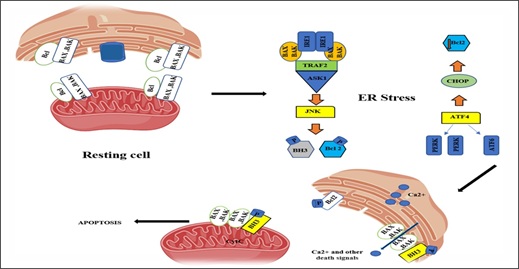

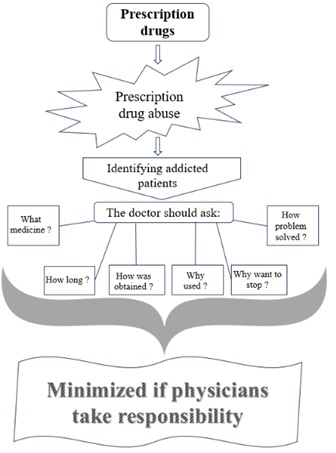
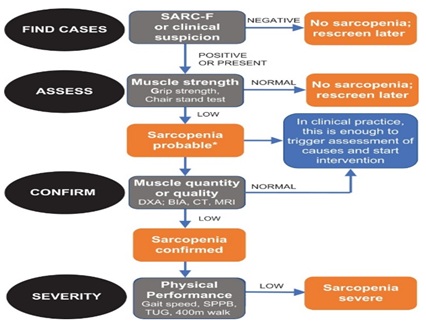
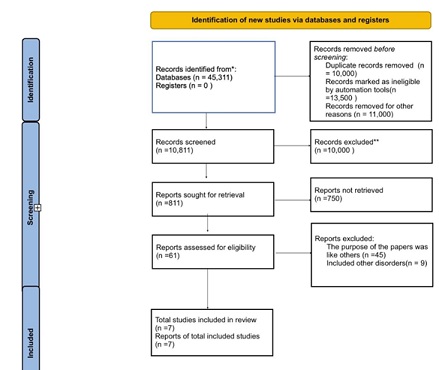


© 2024, Copyrights Herald Scholarly Open Access. All Rights Reserved!